Citoscheletro
Il citoscheletro è una fondamentale struttura cellulare. Il citoscheletro costituisce, per analogia macroscopica, la struttura muscolare, di movimento ed ossea, o di sostegno della cellula. Le sue funzioni di tipo strutturale sono indispensabili quindi in quelle cellule eucariotiche prive di parete, inoltre permette il movimento sia intercellulare (spostamento e modificazioni di forma di tutta la cellula) che intracellulare (spostamento interno dei costituenti la cellula).
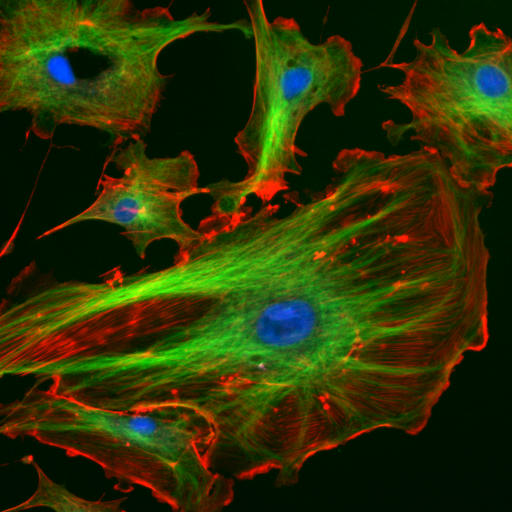
Il citoscheletro è costituito fondamentalmente da tre tipi di filamenti proteici che si distinguono per funzione e composizione:
- filamenti actinici o microfilamenti
- filamenti intermedi
- microtubuli
Filamenti actinici
La componente proteica di questo tipo di filamenti è l'actina. L'actina è una proteina globulare che lega ATP.
Con il loro spessore di 6-7nm, sono i filamenti citoscheletrici più sottili. Hanno una polarità strutturale, ovvero hanno un'estremità con carica elettrica positiva, dove l'aggiunta di g-actina avviene velocemente e contribuisce quindi all'allungamento del filamento, ed una parte con carica elettrica negativa, che influisce poco sull'accrescimento.
La polimerizzazione inizia lentamente con 3 molecole di actina che si legano tra loro. Nella cellula la concentrazione di actina libera è molto alta, quindi altre molecole di actina si legano a questo polimero neoformato ed il processo diventa man mano più veloce fino a che non si giunge ad un punto di equilibrio con l'actina libera nella cellula.
Le proteine associate ai filamenti actinici sono numerose: ci sono proteine che inibiscono la polimerizzazione dei filamenti, altre che tagliano i filamenti e altre ancora che li incappucciano per evitare che si accrescano; ci sono anche proteine che collegano i microfilamenti per formare un fascio, come avviene nei microvilli, ed altre proteine che conferiscono la contrattilità ai filamenti actinici rendendoli così capaci di far cambiare forma alla cellula e di dirigere i traffici interni ad essa.
La miosina
È una proteina presente in tutte le cellule eucariotiche. È dotata di attività ATPasica, cioè è in grado di idrolizzare l'ATP (adenosintrifosfato).
Esistono diverse isoforme di miosina all'interno delle cellule. Nel complesso le varie isoforme funzionano da "motori proteici" per l'actina; in pratica accoppiano l'idrolisi della molecola di ATP con cambiamenti conformazionali che contribuiscono a generare la forza meccanica per i vari tipi di motilità cellulare e sub-cellulare (contrazione cellulare, citocinesi, traffico di vescicole).
La struttura della miosina consiste in due parti principali: la testa globulare, che lega la molecola di actina ed è dotata di attività ATPasica (cioè in grado di idrolizzare l'ATP) e da una coda, unita alla testa, che consiste in due catene proteiche con conformazione ad elica avvolte insieme.
Il complesso actina-miosina, nelle cellule muscolari scheletriche dei Vertebrati, forma una struttura caratteristica detta sarcomero, da cui dipende la contrazione delle fibre muscolari. La contrazione muscolare è influenzata essenzialmente dalla concentrazione intracellulare dello ione calcio, ma anche da altre proteine, come la tropomiosina, la troponina e la nebulina.
Filamenti intermedi
Chiamati così per il loro spessore (circa 10nm) intermedio tra quello dei microtubuli e quello dei filamenti actinici. Le molecole che li costituiscono sono filamentose e variano a seconda del tipo di cellula. Possiedono una grande resistenza alla trazione e consentono alla cellula di sopportare stress meccanici. A differenza degli altri filamenti citoscheoletrici, i filamenti intermedi non sono polarizzati e sono più stabili.
La polimerizzazione dei filamenti intermedi avviene nel seguente modo: 2 molecole di gas si aggregano formando un dimero, che va ad unirsi ad un altro dimero formando un tetramero; infine i tetrameri si aggregano a loro volta fino a che non arrivano a formare un filamento di 32 molecole base molto simile ad una corda.
Una categoria di filamenti intermedi presenti in tutte le cellule è quella delle lamìne, ovvero quel particolare tipo di filamenti che va a costituire la lamina nucleare. Nel processo di divisione cellulare la lamina nucleare deve scomparire, altrimenti il materiale genetico della cellula non potrebbe ripartirsi tra le due cellule figlie. È necessario quindi che le proteine che costituiscono i filamenti intermedi della lamina nucleare vengano fosforilate, in modo da renderle instabili e portarle alla depolimerizzazione. I filamenti intermedi della lamina nucleare sono controllati nei loro processi di polimerizzazione e depolimerizzazione dalla proteina chinasi.
Microtubuli
Sono tubi proteici cavi del diametro di 25nm (diametro esterno) capaci di autodemolirsi rapidamente in una sede e ricostituirsi altrettanto velocemente in un'altra. Le loro pareti sono formate da 13 protofilamenti. Anche i microtubuli sono polari. Sono composti da eterodimeri formati da una molecola di tubulina-α e una di tubulina-β. La tubulina è una proteina capace di legarsi a GTP, ma solo la tubulina-β può idrolizzare GTP a GDP.
Il processo di polimerizzazione dei microtubuli inizia molto lentamente: più eterodimeri di tubulina-αβ, tramite un processo detto di nucleazione, vanno a formare un piccolo microtubulo. Una volta formato, il piccolo microtubulo si accresce da entrambe le parti (la velocità di accrescimento è maggiore all'estremità positiva) ed il completamento è rapido.
In vitro si osserva che, a bassissime concentrazioni di tubulina libera, sia l'estremità positiva che quella negativa si accorciano. Man mano che la concentrazione di tubulina libera aumenta, la depolimerizzazione rallenta fino a che non si raggiunge un punto di equilibrio, detto punto critico; l'equilibrio che si raggiunge è di tipo dinamico. In vitro si può osservare il cosiddetto fenomeno a mulinello: se le concentrazioni di tubulina libera sono abbastanza elevate, i microtubuli si formano spontaneamente.
Nella cellula la concentrazione di tubulina libera non è sufficiente da poter permettere la formazione spontanea dei microtubuli, i quali per formarsi hanno quindi bisogno di un punto di innesco che, nelle cellule animali è dato dalla tubulina-γ, una molecola a forma di anello presente sul centrosoma. Mantenendo bassa la concentrazione di tubulina libera, la cellula può controllare la formazione dei microtubuli, i quali presenteranno quindi un'instabilità dinamica, ovvero si depolimerizzerano e ripolimerizzerano di continuo a partire dal centrosoma. I microtubuli possono stabilizzarsi se alla loro estremità positiva si forma un cappuccio a GTP, che si forma se prima che la tubulina-β idrolizzi il GTP si attacca al microtubulo un altro eterodimero. Un microtubulo che nasce dal centrosoma può non depolimerizzarsi se alla sua estremità positiva (quella più lontana da centrosoma) si va ad attaccare una molecola o una struttura cellulare.
Due microtubuli possono scorrere l'uno sull'altro grazie a speciali proteine presenti sui microtubuli che trasformano l'energia derivante dall'idrolisi di ATP in energia motrice. Ll microtubulo possono attaccarsi anche organuli e vescicole, che possono quindi scorrere per mezzo delle proteine motrici verso l'estremità positiva (le proteine motrici saranno chinesine) o verso l'estremità - (le proteine motrici saranno dineine) del microtubulo.
Ciglia e flagelli
Speciali proteine dette MAPs (microtubules associated proteins) possono stabilizzare in maniera permanente i microtubuli che vanno così a formare ciglia e flagelli.
Le ciglia servono per il movimento della cellula. Generalmente sono numerose sulla superficie cellulare. Possono creare correnti nel liquido intorno alla cellula in modo da indirizzare il cibo verso il luogo in cui verrà digerito, come succede per esempio nelle spugne. La parte interna di un ciglio è detta assonema ed è costituito da una membrana che racchiude 9 coppie di microtubuli alla periferia più 2 microtubuli non accoppiati al centro. Questa struttura è detta 9+2 e si ritrova in quasi tutte le forme di ciglia e flagelli eucariotici, dai protozoi all'uomo. l'assonema si attacca al corpuscolo basale, anch'esso formato da microtubuli, con una struttura leggermente diversa da quella dell'assonema: ci sono 9 triplette ai lati. Nelle ciglia si trovano solo le dineine come proteine motrici.