Homo ergaster
| Homo ergaster | |
|---|---|

| |
| Die skedel KNM-ER 3733, wat in 1975 in Kenia ontdek is. | |
| Wetenskaplike klassifikasie | |
| Koninkryk: | |
| Filum: | |
| Klas: | |
| Orde: | |
| Familie: | |
| Genus: | |
| Spesie: | H. ergaster
|
| Binomiale naam | |
| †Homo ergaster (Colin Groves en Vratislav Mazák, 1975)
| |
Homo ergaster is ’n uitgestorwe spesie van Homo wat tydens die vroeë Pleistoseen in Oos- en Suider-Afrika geleef het.[1] Of H. ergaster 'n spesie op sy eie vorm en of dit in die spesie H. erectus opgeneem moet word, is 'n deurlopende en onopgeloste dispuut tussen paleoantropoloë. Voorstanders van 'n gesamentlike spesie praat gewoonlik van H. ergaster as "Homo erectus uit Afrika"[2] of "Homo erectus ergaster".[3]
Die naam Homo ergaster kan rofweg vertaal word as "werkende man",[1] 'n verwysing na die meer gevorderde gereedskap wat die spesie gebruik het in vergelyking met dié van hulle voorouers. Die fossielomvang van H. ergaster strek van sowat 1,7 miljoen tot 1,4 miljoen jaar gelede, hoewel 'n groter omvang moontlik is.[4]
Hoewel fossiele uit die hele Oos- en Suider-Afrika bekend is, is die meeste H. ergaster-fossiele aan die oewers van die Turkanameer in Kenia ontdek. Daar is latere Afrika-fossiele, waarvan sommige jonger as 'n miljoen jaar is, wat dui op langtermyn- anatomiese kontinuïteit, maar dit is onseker of hulle amptelik beskou kan word as H. ergaster-spesimens. As 'n aaneenlopende spesie kon H. ergaster gelewe het tot so laat as 600 000 jaar gelede, toe nuwe linies van Homo in Afrika ontstaan het.
Dié wat glo H. ergaster moet in H. erectus opgeneem word, reken daar is te min verskille tussen die twee om hulle as aparte spesies te beskou. Voorstanders van twee aparte spesies haal morfologiese verskille aan tussen die fossiele van Afrika en dié van Asië, en reken verder dat vroeë Homo-evolusie veel ingewikkelder is as om die twee spesies as een te beskou. Daarbenewens kan morfologiese verskille tussen spesimens wat almal as H. ergaster beskou word, daarop dui dat H. ergaster self nie 'n samehangende spesie verteenwoordig nie. Ongeag die beste klassifikasie vir hulle, toon H. ergaster primitiewe weergawes van eienskappe wat later in H. erectus voorgekom het en hulle is dus dalk die direkte voorouers van die latere H. erectus-bevolkings in Asië. Hulle is ook waarskynlik die voorouers van die latere hominiene in Europa en Afrika, soos die moderne mense en die Neanderdallers.
Verskeie eienskappe onderskei H. ergaster van Australopithecus, sowel as van vroeëre, meer basale Homo-spesies, soos H. habilis. Daaronder tel groter liggaamsmassa, relatief lang bene, verpligte tweevoetigheid, relatief klein kakebene en tande (wat dui op 'n groot verandering in eetgewoontes), sowel as liggaamsafmetings en lewenstyle wat meer met dié van die moderne mens ooreenstem as met dié van vroeëre of kontemporêre hominiene. Daarom beskou sommige navorsers H. ergaster as die vroegste ware verteenwoordiger van die genus Homo. Richard Dawkins het dit verkies en bygevoeg: "Ek sal hulle Ergasts eerder as Erects noem, deels omdat ek glo die meeste van ons gene kom van die Afrika-vorm en deels omdat hulle nie enigsins meer regop as hulle voorouers was nie".[5]
H. ergaster het op die savanne van Afrika gewoon, 'n unieke omgewing wat tot die behoefte aan nuwe en unieke gedrag kon gelei het. Vroeë Homo-lede het waarskynlik, nes moderne primate, van teenaanvalstegnieke gebruik gemaak om roofdiere weg te hou. Teen die tyd van H. ergaster het dié gedrag moontlik gelei tot die ontwikkeling van ware jagter-versamelaarsgedrag, 'n eerste onder primate. H. ergaster was 'n toproofdier.[6] Nog gedrag wat eerste in dié spesie kon begin het, sluit in die verdeling van mans en vroue in kosversameling en ware monogame bande tussen pare. H. ergaster het ook gevorderder Acheuleense gereedskap gebruik, onder meer die eerste bekende handbyle. Hoewel onbetwiste bewyse ontbreek, kon H. ergaster ook die eerste hominien gewees wat vuur beheer het.
Taksonomie
[wysig | wysig bron]Navorsingsgeskiedenis
[wysig | wysig bron]
Die sistematiek en taksonomie van Homo in die vroeë tot middelste Pleistoseen is een van die mees betwiste gebiede van paleoantropologie.[7] In vroeë paleoantropologie en tot laat in die 20ste eeu is algemeen aanvaar Homo sapiens is die eindresultaat van geleidelike veranderings binne 'n enkele linie van die hominien se evolusie. Toe al hoe meer fossiele van die waargenome oorgangsvorm tussen vroeë hominiene en moderne mense, H. erectus, wat aanvanklik beskou is as argaïese menslike fossiele in Asië, ontdek word, het die uiteenlopendheid binne H. erectus daaroe gelei dat sommige mense begin vra het wat die spesie definieer en wat die omvang daarvan moet wees. Sommige navorsers, soos die paleoantropolooog Ian Tattersall het H. erectus in 2013 bevraagteken omdat dit 'n ongelooflike getal fossiele behels het met "aansienlik verskillende vorme".[8]
In die 1970's het Richard Leakey en Alan Walker 'n reeks hominienfossiele aan die oostelike oewer van die Turkanameer in Kenia beskryf. Die merkwaardigste vondste was twee gedeeltelike skedels: KNM ER 3733 en KNM ER 3883, wat by Koobi Fora gevind is. Leakey en Walker het die skedels aan H. erectus toegeskryf en aangeteken dat hulle breinvolumes (onderskeidelik 848 en 803 cm3) goed vergelyk met dié van die veel jonger tipespesimen van H. erectus (950 cm3). Nog 'n belangrike fossiel was 'n fossiele kaak met die naam KNM ER 992 wat in 1972 by Ileret ontdek is en deur Leaky beskryf is as "Homo van onbepaalde spesie".[9]

In 1975 het Colin Groves en Vratislav Mazák die fossiel KNM ER 992 beskryf as die holotipespesimen van 'n aparte spesie, wat hulle Homo ergaster genoem het.[10] Die naam "ergaster" is afgelei van die antieke Griekse ἐργαστήρ (ergastḗr, "werker" of "werksman").[11][12] Groves en Mazák het ook baie van die Koobi Fora-fossiele, soos KNM ER 803 ('n gedeeltelike skelet en 'n paar geïsoleerde tande) ingesluit in die nuwe spesie, maar het geen vergelyking met die Asiatiese fossielrekord van H. erectus in hulle beskrywing ingesluit nie. Dit het 'n deel van die taksonomiese verwarring oor die spesie veroorsaak.[13]
'n Byna volledige fossiel, wat as 'n jong man beskou is (hoewel die geslag eintlik nooit vasgestel is nie) is in 1984 deur die Keniaanse argeoloog Kamoya Kimeu aan die westelike oewer van die Turkanameer ontdek.[12] Leakey en Walker, saam met Frank Brown en John Harris, het dit in 1985 beskryf as KNM-WT 15000 (met die bynaam "Turkana-seun"). Hulle het die feitlik volledige skelet as lid van H. erectus beskou.[14] Die Turkana-seun was die eerste goed bewaarde spesimen van H. ergaster/erectus wat ontdek is en is 'n belangrike fossiel in die vasstelling van die verskille en ooreenkomste tussen vroeë Homo en moderne mense.[15] Die paleoantropoloog Bernard Wood het die Turkana-seun in 1992 in H. ergaster geplaas.[12] Dit word vandag, saam met ander fossiele in Afrika, wat vroeër as H. erectus gereken is, algemeen beskou as verteenwoordigend van H. ergaster deur diegene wat die twee as aparte spesies beskou.[16]
Klassifikasie
[wysig | wysig bron]H. ergaster word maklik van vroeëre en meer basale spesies van Homo, veral H. habilis en H. rudolfensis, onderskei deurdat verskeie eienskappe en hulle lewenstyl nader aan dié van die moderne mens was as aan vroeëre en kontemporêre hominiene. In vergelyking met hulle verwante, was H. ergaster se liggaamsproporsies meer soortgelyk aan die latere lede van die genus Homo. Hulle het relatief lang bene gehad, wat hulle verpligtend tweevoetig gemaak het. Hulle tande en kake was ook kleiner as dié van H. habilis en H. rudolfensis, wat dui op 'n groot verandering in eetgewoontes.[17] In 1999 het die paleoantropoloë Bernard Wood en Mark Collard gesê die konvensionele kriteria wat gebruik word om spesies aan die genus Homo toe te ken, is foutief en dat vroeë en basale spesies soos H. habilis en H. rudolfensis dalk herklassifiseer moet word as voorouers van Australopithecus. Volgens hulle was H. ergaster die eerste ware verteenwoordiger van Homo.[18]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Kladogram deur Strait, Grine & Fleagle (2015)[19] |
Sedert die beskrywing van H. ergaster as 'n aparte spesie in 1975, word die klassifikasie van fossiele daarin betwis. H. ergaster is dadelik verwerp deur Leakey en Walker en baie ander invloedryke navorsers, soos die paleoantropoloog G. Philip Rightmire, wat in 1990 'n uitgebreide verhandeling oor H. erectus geskryf het. Hy het 'n inklusiewer H. erectus verkies.
Oor die algemeen is daar min twyfel dat die groep fossiele wat H. erectus en H. ergaster uitmaak, die fossiele van 'n min of meer samehangende stel nabye verwante van argaïse mense verteenwoordig. Die vraag is eerder of hierdie fossiele verskillende spesies verteenwoordig of 'n enkele, hoogs uiteenlopende spesie oor die verloop van byna twee miljoen jaar.[10] Dié deurlopende debat bly onopgelos. Navorsers gebruik gewoonlik die term "Homo erectus sensu stricto" vir die Asiatiese H. erectus en "Homo erectus sensu lato" vir die groter spesie wat beide die Afrika-weergawe (H. ergaster) en die Asiatiese weergawe insluit.[20][21]


Om vanselfsprekende redes deel H. ergaster baie eienskappe met H. erectus, soos lang, vooruitstekende kakebene, groot wenkbrouriwwe en 'n kop wat skuins na agter loop.[22] Baie eienskappe van H. ergaster is duidelik primitiewer weergawes van eienskappe wat later in H. erectus voorgekom het, en dit verduister die verskille tussen die twee effens.[23] Daar is subtiele, moontlik belangrike, verskille tussen die fossiele van Oos-Afrika en dié van Oos-Asië. Twee daarvan is H. ergaster se kopbene met 'n ietwat hoër koepel en dunner wande, en die selfs groter wenkbrouriwwe en gesigte van H. erectus.[22]
Die vraag word bemoeilik omdat dit betref hoe groot die verskille in 'n enkele spesie kan wees voordat dit in meer spesies verdeel moet word, en daarvoor is daar nie 'n duidelike antwoord nie. In 'n ontleding in 2008 deur die antropoloog Karen L. Baab van fossiele van verskeie H. erectus-subspesies, asook ander wat in H. ergaster val, is bevind die intraspesifieke verskille in H. erectus is groter was wat verwag kan word vir 'n enkele spesie wanneer dit vergelyk word met moderne mense en sjimpansees. As dit met gorillas vergelyk word, val dit binne die verwagte variasie, en ook binne die omvang van 'n enkele subspesie wanneer dit met die orangoetang vergelyk word (hoewel dit deels vanweë die groot geslagsdimorfie in gorillas en orangoetangs is).[24] Baab het tot die gevolgtrekking gekom H. erectus s.l. is óf 'n enkele, maar veranderlike spesie óf verskeie subspesies wat deur tyd en geografie verdeel is óf verskeie geografies verstrooide, maar nabyverwante spesies.[25]
In 2015 het die paleoantropoloë David Strait, Frederick Grine en John Fleagle H. ergaster gelys as een van sewe "algemeen erkende" spesies van Homo, saam met H. habilis, H. rudolfensis, H. erectus, H. heidelbergensis, H. neanderthalensis en H. sapiens. Hulle het bygevoeg ander spesies, soos H. floresiensis en H. antecessor, is minder bekend.[19]
Variasie in die fossielmateriaal
[wysig | wysig bron]



Toe Ian Tattersall in 2013 verskeie fossiele van H. ergaster vergelyk met dié van H. erectus, veral die tipespesimen, het hy tot die gevolgtrekking gekom om na die materiaal in Afrika as H. ergaster te verwys eerder as na die "H. erectus uit Afrika", is 'n "aansienlike verbetering", omdat daar baie eienskappe is wat die materiaal van die twee kontinente onderskei.[26] Tattersall het geglo dit is passend om die term H. erectus net vir Oos-Asiatiese fossiele te gebruik in plaas van vir fossiele in Asië én Afrika. Hoewel hy gereken het die H. ergaster-materiaal verteenwoordig die fossiele van 'n aparte klade van Homo, het hy ook bevind daar is aansienlike diversiteit binne die klade: KNM ER 992 se kaak stem goed ooreen met ander fossiele kake uit die streek, soos dié van OH 22 van Olduvai en KNM ER 3724 van Koobi Fora, maar stem nie noodwendig ooreen met die skedels nie, soos dié van KNM ER 3733 en KNM ER 3883, en ook nie met die kaak van die Turkana-seun nie omdat die tande grootliks verskil.[26]
Die "ikoniesste" fossiel van H. ergaster is die skedel KNM ER 3733, wat deur 'n paar eienskappe skerp van die Asiatiese H. erectus onderskei word, insluitende dat die wenkbrouriwwe boontoe én vorentoe uitsteek en dat die harsingskas taamlik lank in vergelyking met sy breedte is, met die kantwande wat 'n boog maak. KNM ER 3733 kan ook van KNM ER 3883 onderskei word deur 'n paar eienskappe, veral dat die kante van KNM ER 3883 se wenkbrouriwwe baie verdik is en wel vorentoe uitsteek, maar effens na onder en nie na bo nie.[27] Albei skedels verskil van dié van die Turkana-seun, wat net effens verdik is, maar nie vertikaal verdik is soos KNM ER 3883 s'n of baie ver vorentoe uitsteek soos KNM ER 3733 s'n nie. Daarby is die Turkana-seun se gesigstruktuur smaller en langer as die ander skedels. Dit het ook hoër neusopenings en waarskynlik 'n platter profiel van die boonste deel van die gesig. Dit kan wees omdat hy net tussen sewe en 12 jaar oud was.[28] Verder is die veronderstelling dat KNM ER 3733 die skedel van 'n vrou was en die Turkana-seun aanvanklik vermoedelik manlik was, wat beteken geslagsdimorfie kan vir van die verskille verantwoordelik wees.[15]
Die verskille tussen die Turkana-seun se skedel en beide KNM ER 3733 en KNM ER 3883, sowel as die verskille in die tande van die seun en KNM ER 992 is al deur sommiges, soos die paleoantropoloog Jeffrey H. Schwartz, vertolk as dat die Turkana-seun en die res van die H. ergaster-materiaal nie noodwendig dieselfde takson verteenwoordig nie. Schwartz het ook opgemerk dat nie een van die fossiele H. erectus verteenwoordig nie en dat die materiaal aansienlik hersien moet word.[29] In 2000 het die Franse paleoantropoloog Valéry Zeitoun voorgestel daar moet na KNM ER 3733 en KNM ER 3883 as twee aparte spesies verwys word, wat hy H. kenyaensis (tipespesimen KNM ER 3733) en H. okotensis (tipespesimen KNM ER 3883) genoem het, maar dié name het nie algemeen byval gevind nie.[30]
Evolusionêre geskiedenis
[wysig | wysig bron]Evolusie en tydsomvang
[wysig | wysig bron]Hoewel dikwels aanvaar word dat H. ergaster in Oos-Afrika ontstaan het, word hulle oorsprong verduister deurdat die spesie soveel anders as vroeëre spesies van Homo en Australopithecus was; hulle het lang ledemate en moderne liggaamsverhoudings gehad en was lank. Baie gereedskap uit die Pleistoseen is in Oos-Afrika ontdek, maar dit is nie heeltemal seker dat H. ergaster se oorsprong daar lê sonder nog fossielvondste nie.[31] Daar word aanvaar H. ergaster het ontwikkel uit vroeëre Homo-spesies, moontlik H. habilis. Fossiele wat ontdek is, was hoofsaaklik van Oos-Afrika in die tydsomvang 1,8 miljoen tot 1,7 miljoen jaar gelede. Die meeste is aan die oewers van die Turkanameer in Kenia gevind.[11]
Die oudste bekende spesimen van H. ergaster is DNH 134, 'n skedel wat in die Drimolen-grotstelsel in Suid-Afrika ontdek is; dit was van 2,04 miljoen tot 1,95 miljoen jaar gelede. Die skedel is ook die oudste bekende H. erectus s.l.-spesimen en wys groot ooreenkomste met KNM ER 3733. Dit wys ook vroeë H. ergaster het saam met ander hominiene soos Paranthropus robustus en Australopithecus sediba bestaan.[32]
Daar is ook jonger spesimens van H. ergaster; die Turkana-seun dateer van sowat 1,56 miljoen jaar gelede.[11] 'n Paar selfs jonger Afrika-skedels dui op langtermyn- anatomiese kontinuïteit, maar dit is onduidelik of hulle werklik as H. ergaster-spesimens beskou kan word; die "Olduvai-hominied 9"-skedel van die Olduvai-ravyn wod gedateer van sowat 1,2 miljoen tot 1,1 miljoen jaar gelede. Daar is ook skedels van Buia (naby die kus van Eritrea, sowat 1 miljoen jaar oud), die Bouri-formasie in Ethiopië (sowat 1 miljoen tot 780 000 jaar oud) en 'n gedeeltelike skedel van Olorgesailie in Kenia (tussen 970 000 en 900 000 jaar oud). Die Olduvai-skedel is soortgelyk aan dié van die Asiatiese H. erectus deurdat hy swaar wenkbrouriwwe het, maar die ander verskil net effens van vroeëre H. ergaster-skedels.[33]
Die H. erectus in Asië, sowel as latere hominiene in Europa (soos H. heidelbergensis en H. neanderthalensis) en Afrika (H. sapiens) stam moontlik almal van H. ergaster af.[11] Omdat H. ergaster vermoedelik die voorouers van hierdie latere Homo-spesies was, kon dit in Afrika bestaan het tot omtrent 600 000 jaar gelede, toe breingrootte vinnig toegeneem en H. heidelbergensis ontstaan het.[34]
Verspreiding uit Afrika
[wysig | wysig bron]- Sien ook: Uit Afrika-teorie
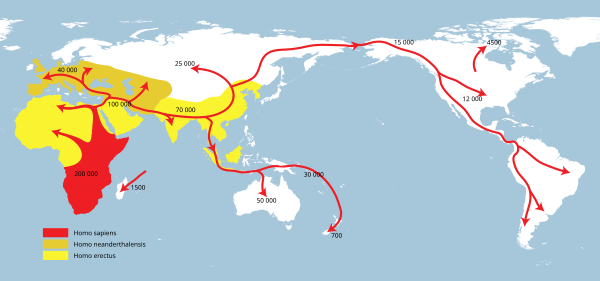
H. erectus word tradisioneel beskou as die eerste hominien wat Afrika verlaat het om in Europa en Asië te gaan woon. As H. ergaster 'n aparte spesie van H. erectus is, sal dié rol eerder op H. ergaster toegepas kan word.[26][11] Baie min konkrete inligting bestaan oor wanneer en watter Homo-spesie eerste in Europa en Asië aangekom het, want fossiele uit die vroeë Pleistoseen is op albei kontinente skaars. Die teenwoordigheid van H. erectus-fossiele in Oos-Asië beteken 'n menslike spesie, heel waarskynlik H. ergaster, het Afrika meer as 1 miljoen jaar gelede verlaat.[35] Vroeër is gereken hulle is tussen 1,9 miljoen en 1,7 miljoen jaar gelede uit Afrika uit.[22] Ontdekkings in Georgië en China stoot die laaste moontlike datum verder terug: tot voor 2 miljoen jaar gelede. Dit werp ook twyfel op die idee dat H. ergaster die eerste hominien was wat Afrika verlaat het.[36]
Die algemene aanvaarding is dat hominiene óf oor die suidekant van die Rooisee óf met die Nylvallei langs uit Afrika migreer het, maar daar is nie in een van dié streke enige fossiele hominiene uit die vroeë Pleistoseen nie. Die vroegste bekende Homo-fossiele buite Afrika is die Dmanisi-skedels van Georgië (tussen 1,77 miljoen en 1,85 miljoen jaar oud),[36] wat óf 'n vroeë H. ergaster óf 'n nuwe takson (H. georgicus) verteenwoordig; die tande van Ubeidiya in Israel (sowat 1,4 miljoen tot 1 miljoen jaar oud) en die fossiele van die Javamens (H. erectus erectus, meer as 8 000 km van daar).[31] Ubeidiya is ook die oudste bevestigde vindplek buite Afrika van Acheuleense gereedskap (iets wat met H. ergaster verbind word). Die gereedskap wat ontdek is, lyk baie soos ouer gereedskap wat in Oos-Afrika gevind is.[35]
Die vroegste fossielbewyse van Homo in Asië is die genoemde Dmanisi-skedels, wat baie eienskappe met H. ergaster in Afrika deel. Dit dui daarop dat H. ergaster so vroeg as 1,7 miljoen tot 1,9 miljoen jaar gelede uit Afrika versprei het.[37] Benewens eienskappe van H. ergaster, stem baie van die Dmanisi-skedels se eienskappe ook ooreen met dié van vroeëre hominiene, soos H. habilis, en die terrein ontbreek bewaarde handbyle (wat met H. ergaster verbind word). Dit beteken hominiene kon selfs voor H. ergaster uit Afrika versprei het.[21] Veral skedel D2700 (Dmanisiskedel 3) lyk soos dié van H. habilis. Dit beteken meer as twee groepe argaïese mense kon Afrika vroeg verlaat het.[33] Verder is steengereedskap wat deur hominiene gemaak is, op die Loessplato China ontdek en gedateer as 2,12 miljoen jaar oud, wat beteken hominiene moes Afrika voor dié tyd verlaat het.[36]
'n Alternatiewe historiese hipotese is dat Homo in Asië ontstaan het uit vroeëre voorouers wat uit Afrika daarheen migreer het. Hulle het terug na Europa versprei, waar dit gelei het tot H. sapiens. Dit was die mening van Eugène Dubois, wat die Javamens in die 19de eeu beskryf het wat toe die oudste bekende hominienfossiele was. Hy het dit gebruik om die hipotese te bewys. Hoewel die ontdekking van Australopithecus en vroeëre Homo in Afrika beteken Homo self se oorsprong lê nie in Asië nie, word die idee dat H. erectus (of H. ergaster) wel het, en toe terug na Afrika versprei het, soms weer opgehaal.[38]
Anatomie
[wysig | wysig bron]Bou en voorkoms
[wysig | wysig bron]
Die enigste goed bewaarde skeletfossiel van H. ergaster is dié van die Turkana-seun. Anders as Australopithecus, was die verhouding tussen die seun se arms en bene nie groter as dié van lewende mense nie. Verder het die keëlvormige torso van hulle voorouers ontwikkel in 'n meer vaatjievormige bors bo smal heupe, nog 'n ooreenkoms met moderne mense.[39] Die skeenbeen van die Turkana-seun is relatief langer as dié in mense, wat dalk beteken hulle knieë het meer gebuig wanneer hulle geloop het.[40] Die skraal, lang bou van die Turkana-seun kan verduidelik word aan die hand daarvan dat H. ergaster in warm, droë omgewings gewoon het, want 'n skraal liggaam beteken groter hitteverspreiding.[41]
H. ergaster-individue was aansienlik langer as hulle voorouers. Terwyl die beroemde Australopithecus-fosiel Lucy sowat 'n meter lank sou gewees het met haar dood, was die Turkana-seun sowat 1,62 m lank, en hy sou moontlik as 'n volwassene tot 1,82 m lank geword het.[39]
Omdat H. ergaster by 'n warm, droë klimaat aangepas het, was hulle dalk die eerste menslike spesie wat 'n haarlose liggaam gehad het.[41] Hoewel sweet die algemeen aanvaarde verduideliking vir haarloosheid is, is ander moontlike redes minder parasiete[42] en seksuele seleksie.[43]
Haarverlies kon ook vroeër as by H. ergaster voorgekom het. Hoewel geen hominienfossiele velafdrukke gelaat het nie, is dit moontlik dat menslike voorsate reeds drie miljoen jaar gelede hare verloor het. Menslike voorouers het sowat drie miljoen jaar gelede platluise van gorillas gekry en spesievorming van gorilla- na menslike platluise was dalk net moontlik omdat mense teen dié datum die meeste van hulle liggaamshare verloor het.[44]
Skedel en gesig
[wysig | wysig bron]
Verskille met moderne mense sou geredelik sigbaar gewees het in die gesigte en skedels van H. ergaster. Die Turkana-seun was amper volgroei tot hy dood is, maar die volume van sy skedel (880 cm3) was net sowat 130 cm3 groter as die maksimum van H. habilis, wat sowat 500 cm3 onder die mens se gemiddelde was. Die toename van 130 cm3 van H. habilis af raak heelwat kleiner as 'n mens die liggaamsgrootte van die Turkana-seun en H. ergaster in ag neem.[45] Met al die skedels van H. ergaster in ag genome, wissel die spesie se breinvolume gewoonlik tussen 600 en 910 cm3. Omdat hulle breine kleiner as mense s'n was, het hulle skedels dadelik agter die oogkaste vernou.[40]

Die harsingskas was lank en laag, en die Turkana-seun se voorkop was plat. 'n Merkbare verskil tussen die Turkana-seun aan die een kant en Australopithecus en H. habilis aan die ander, sou eersgenoemde se neus gewees het. Dit sou baie soos die mens s'n gelyk het: Dit het vooruit gesteek en die neusgate was afwaarts gedraai. Dié eksterne neus kon ook 'n aanpassing by 'n warmer klimaat gewees het, want mense se neus is gewoonlik koeler as die res van hulle liggaam. Dit kondenseer klammigheid wat andersins uitgeasem sou geword en verlore sou gegaan het tydens tydperke van groter aktiwiteit.[45] Die Turkana-seun se gesig sou langer as die mens s'n gewees het, en die kakebeen sou verder vooruit gesteek het. Hoewel die kakebene en tande kleiner as dié van die gemiddelde Australopithecus en H. habilis was, was hulle steeds heelwat groter as moderne mense s'n.[41]
Hoewel die Turkana-seun ouer as 12 jaar was toe hy dood is, stem sy gestalte meer ooreen met 'n moderne 15-jarige s'n en sy brein met 'n moderne eenjarige s'n. Teen moderne standaarde sou H. ergaster dus kognitief beperk gewees het, hoewel die uitvinding van nuwe gereedskap wys hulle was intelligenter as hulle voorouers.[46]
Liggaamsmassa en geslagsdimorfie
[wysig | wysig bron]
H. ergaster het 'n aansienlik groter liggaamsmassa gehad as vroeëre hominiene soos die vroeë Homo, Australopithecus en Paranthropus.[17] Australopithecus se gewig het gewoonlik gewissel tussen 29 en 48 kg en H. ergaster s'n tussen 52 en 63 kg.[47] Dit is moontlik dat die groter liggaam die resultaat van 'n lewe in die oop savanne was, omdat meer kos in groter gebiede gesoek en groter prooi gejag kon word, en omdat dit beweeglikheid vergroot het.[17] Die groter liggaamsmassa sou ook beteken het ouers sou hulle kinders kon dra totdat hulle ouer en swaarder was.[47]
Hoewel geslagsdimorfie histories genoem is as een van die groot verskille tussen H. ergaster en vroeëre Homo en Australopithecus,[17][31] is dit moeilik om geslagsdimorfie in uitgestorwe spesies te bepaal, omdat die geslag van die fossiele gewoonlik nie vasgestel kan word nie.
Groei en ontwikkeling
[wysig | wysig bron]Die dimensie van 'n 1,8 miljoen jaar oue, volwasse, vroulike H. ergaster-bekken van Gona, Ethiopië, dui daarop dat H. ergaster in staat sou gewees het om die lewe te skenk aan kinders met 'n maksimum voorgeboortelike breingrootte van 315 cm3, sowat 30-50% van 'n volwassene se brein. Dit val sowat halfpad tussen sjimansees (~40%) en moderne mense (28%).[48] Die Mojokerto-kind, 'n eenjarige Asiatiese H. erectus van tussen 1,4 miljoen tot 1,5 miljoen jaar oud, het die breingrootte van sowat 72-84% van dié van 'n volwasse H. erectus gehad, wat dui op 'n breingroei wat meer met dié van ander groot ape ooreenstem as met moderne mense s'n.[49] Beide die Gona-bekken en Mojokerto-kind dui daarop dat die voorgeboortelike groei van H. ergaster soortgelyk aan dié van die moderne mens was, maar die nageboortelike groei en ontwikkeling halfpad tussen sjimansees en moderne mense s'n.[48]
Die vinniger ontwikkelingstempo dui daarop dat 'n langer kindertyd en 'n lang tydperk van afhanklikheid van die ouers in 'n latere stadium van die menslike evolusie ontwikkel het, moontlik in die laaste gemeenskaplike voorsaat van Neanderdallers en moderne mense.[49] Dit kan ook beteken die verwagte lewensduur van H. ergaster en H. erectus was korter as dié van latere en moderne mense.[50]
Kultuur
[wysig | wysig bron]
Eetgewoontes
[wysig | wysig bron]H. ergaster het waarskynlik meer diervet as sy voorgangers geëet. Dit het meer energie toegelaat vir breingroei en het die brein se grootte laat toeneem.[51][52]
As hulle meer energie nodig gehad het, sou H. ergaster baie meer moes geëet het as Australopithecus of sou hulle kos van veel beter gehalte moes geëet het. Die kakebene en tande van H. ergaster was kleiner as dié van Australopithecus, wat dui op 'n verskuiwing in dieet weg van veselryker kos wat moeiliker was om te kou. Hulle het ook 'n klein maag gehad, wat beteken H. ergaster sou kos moes geëet het wat makliker verteerbaar was, wat ook op 'n beter gehalte dui.[53]
H. ergaster het waarskynlik meer vleis as Australopithecus geëet.[53] Vleis is dalk bekom met 'n kombinasie van lokvalle, aktiewe jagmetodes en mededinging om aas. Vleis alleen kon egter nie voldoende gewees het vir die H. ergaster se kosbehoeftes nie. Die topprooi wat beskikbaar was, het min vet gehad, en diëte met baie vleis verhoog die behoefte aan die inneem van water, wat moeilik sou gewees het in 'n oop, warm omgewing.
H. ergaster sou daarom benewens vleis ook 'n groot verskeidenheid ander kosse moes geëet het, soos sade, heuning, neute en ongewerweldes,[54] asook voedsame wortels, knolle en ander ondergrondse plantbergingsorgane.[41] Vanweë hulle klein kouvermoë sou die gebruik van gereedskap nodig gewees het om die kos te bewerk voordat dit geëet is.[55]
Sosiale struktuur en dinamika
[wysig | wysig bron]
H. ergaster het op die Afrika-savanne gewoon, wat gedurende die Pleistoseen die tuiste van veel meer karnivore was as tans. Hominiene kon dalk net by die savanne aangepas het as hulle roofdiere doeltreffend afgeweer het. Dit sou beteken hulle moes in groot groepe gewoon, steen- en dalk houtgereedskap gehad en doeltreffende teenaanvalstrategieë gevestig het. Moderne primate wat baie tyd op die savanne deurbring, soos sjimpansees en bobbejane, vorm groot groepe met baie mannetjies waarin die mannetjies doeltreffend saamwerk om roofdiere af te weer, soms met die gebruik van klippe en stokke, om die groep te beskerm. Dit is moontlik dat die vroeë Homo dieselfde gedrag geopenbaar het. Hulle kon dalk nes bonobo's en sjimpansees manlike bonde gevorm het. Weens die skaarste aan fossiele, is dit moeilik om vas te stel hoe groot die groepe van vroeë Homo was. Groepe was moontlik groot, sowat 100 individue of meer, as na groepe sjimpansees en bobbejane gekyk word.[56]
In H. ergaster het dié gedrag waarskynlik verder ontwikkel. Hulle was moontlik die eerste primate wat jagter-versamelaars was.[56] Predasie (die verorbering van diere in groot groepe) het dalk 'n kaskade van evolusionêre veranderings teweeggebring wat die gang van die menslike ontwikkeling verander het. Koöperatiewe gedag soos jag in groot groepe, verdediging teen roofdiere en mededinging om aas sou kritiek gewees het vir oorlewing, en dus het 'n basiese oorgang in sielkunde moontlik geleidelik plaasgevind. Nes by moderne primate, was jag by argaïese mense vermoedelik hoofsaaklik 'n manlike aktiwiteit. Vroue was waarskynlik verantwoordelik vir die insameling van ander soorte kos, soos vrugte, neute en eiers.[57]
Die deel van kos het waarskynlik hegte bande tussen die lede van 'n groep gesmee. Dit kon gelei het tot vriendskappe tussen mans en vroue en later tot die vorming van monogame paartjies. Omdat vroue in seksuele seleksie 'n voorkeur toon vir mans wat jag, is dié gedrag deur geslagte oorgedra en versterk.[57]
Die enigste regstreekse bewys dat H. ergaster in groepe gewoon het, is 'n paar terreine by Ileret in Kenia, waar 97 voetspore van 1,5 miljoen jaar gelede deur 'n groep van minstens 20 individue bewaar gebly het. Na aanleiding van die grootte van die spore, is een stel net deur mans getrap. Dit dui op 'n gespesialiseerde taak, soos jag, 'n grenspatrollie of 'n soektog na aas. Dit versterk die idee van 'n skeiding tussen mans en vroue wat verantwoordelikhede betref.[58]
Gereedskap
[wysig | wysig bron]- Sien ook: Acheuleen

Vroeë H. ergaster het die Oldowan-gereedskapkultuur van Australopithecus en vroeëre Homo geërf, maar het gou geleer hoe om veel groter steenvlokkies af te skilfer as hulle voorouers en tydgenote. Teen 1,65 miljoen jaar gelede het hulle die uitvoerig afgeskilferde artefakte en vroeë handbyle gemaak wat die Acheuleense kultuur kenmerk,[11] en teen 1,6 miljoen tot 1,4 miljoen jaar gelede was die nuwe gereedskapnywerheid goed in Oos-Afrika gevestig.[59]
Die Acheuleense gereedskap verskil van dié van vroeëre beskawings, wat waarskynlik meestal gebruik is om die doppe van neute mee oop te kraak. Die meeste handbyle van H. ergaster is gemaak om smal, skerp voorwerpe mee te splinter, meestal in 'n traandruppel-, ovaal of driehoekige vorm.[34]
Ondanks die naam het handbyle nie 'n spesifike vorm of funksie gehad nie. Dit het in alle groottes en vorms voorgekom en is vir verskeie doeleindes gebruik. Versigtig gevormde en simmetriese handbyle is dalk gebruik om prooi mee te gooi, terwyl ander bloot gedra is om klippe mee te splinter of hout mee te kap. Dit is dalk ook soms gebruik om karkasse mee af te slag.[60] Die gereedskap het nie veel verander deur die bestaan van H. ergaster nie.[60]
Verwysings
[wysig | wysig bron]- ↑ 1,0 1,1 Hazarika, Manji (16–30 Junie 2007). "Homo erectus/ergaster and Out of Africa: Recent Developments in Paleoanthropology and Prehistoric Archaeology".
- ↑ Tattersall 2013, p. 5.
- ↑ Tattersall 2013, p. 14.
- ↑ Wood, Bernard; Doherty, Dandy; Boyle, Eve (29 Mei 2020). "Hominin Taxic Diversity". Oxford Research Encyclopedia of Anthropology (in Engels). doi:10.1093/acrefore/9780190854584.013.194. ISBN 9780190854584. Besoek op 19 April 2021.
- ↑ R. Dawkins, The Ancestor's Tale, 2004, p.59.
- ↑ Ben-Dor, Miki; Sirtoli, Raphael; Barkai, Ran (2021). "The evolution of the human trophic level during the Pleistocene". American Journal of Physical Anthropology. 175: 27–56. doi:10.1002/ajpa.24247. PMID 33675083.
- ↑ Tattersall 2013, p. 1.
- ↑ Tattersall 2013, p. 2.
- ↑ Tattersall 2013, pp. 3–4.
- ↑ 10,0 10,1 Tattersall 2013, p. 4.
- ↑ 11,0 11,1 11,2 11,3 11,4 11,5 Klein 2005, p. 85.
- ↑ 12,0 12,1 12,2 Roberts 2018, p. 116.
- ↑ Antón 2003, p. 127.
- ↑ Brown et al. 1985, p. 788.
- ↑ 15,0 15,1 Brown et al. 1985, p. 789.
- ↑ Wang et al. 2004, p. 453.
- ↑ 17,0 17,1 17,2 17,3 Aiello & Wells 2002, p. 324.
- ↑ Wood & Collard 1999, p. 65.
- ↑ 19,0 19,1 Strait, Grine & Fleagle 2015, p. 2006.
- ↑ Antón, S. C. (2003), Natural history of Homo erectus. Am. J. Phys. Anthropol., 122: 126–170. doi:10.1002/ajpa.10399
- ↑ 21,0 21,1 Dennell & Roebroeks 2005, p. 1100.
- ↑ 22,0 22,1 22,2 Klein 2005, p. 92.
- ↑ Antón 2003, p. 154.
- ↑ Baab 2008, p. 841.
- ↑ Baab 2008, p. 842.
- ↑ 26,0 26,1 26,2 Tattersall 2013, p. 15.
- ↑ Tattersall 2013, p. 8.
- ↑ Tattersall 2013, p. 9.
- ↑ Schwartz 2000, pp. 55–56.
- ↑ Bonde 2012, p. 171.
- ↑ 31,0 31,1 31,2 Dennell & Roebroeks 2005, p. 1099.
- ↑ Herries et al. 2020.
- ↑ 33,0 33,1 Klein 2005, p. 104.
- ↑ 34,0 34,1 Klein 2005, p. 93.
- ↑ 35,0 35,1 Klein 2005, p. 101.
- ↑ 36,0 36,1 36,2 Zhu et al. 2018, p. 608.
- ↑ Klein 2005, p. 103.
- ↑ Larick & Ciochon 1996, p. 1.
- ↑ 39,0 39,1 Klein 2005, p. 87.
- ↑ 40,0 40,1 Roberts 2018, p. 117.
- ↑ 41,0 41,1 41,2 41,3 Klein 2005, p. 88.
- ↑ Pagel & Bodmer 2004, p. 329.
- ↑ Giles 2010, p. 326.
- ↑ Ruxton & Wilkinson 2011, p. 20967.
- ↑ 45,0 45,1 Klein 2005, pp. 87–88.
- ↑ Klein 2005, p. 89.
- ↑ 47,0 47,1 Aiello & Wells 2002, p. 325.
- ↑ 48,0 48,1 Simpson et al. 2008, p. 1090.
- ↑ 49,0 49,1 Coqueugniot et al. 2004, pp. 299–302.
- ↑ Caspari & Lee 2004.
- ↑ Ungar, Grine & Teaford 2008, pp. 208–228.
- ↑ Ben-Dor et al. 2011.
- ↑ 53,0 53,1 Aiello & Wells 2002, p. 326.
- ↑ Aiello & Wells 2002, p. 327.
- ↑ Janssen, Sept & Griffith 2007, p. 308.
- ↑ 56,0 56,1 Willems & Schaik 2017, p. 17.
- ↑ 57,0 57,1 Willems & Schaik 2017, p. 19.
- ↑ Hatala et al. 2016, pp. 1–7.
- ↑ Klein 2005, p. 94.
- ↑ 60,0 60,1 Klein 2005, p. 95.
Bronne
[wysig | wysig bron]- Deacon, Terrence W. (1998). The Symbolic Species: The Co-evolution of Language and the Brain. W.W. Norton & Company. ISBN 0-393-03838-6.
- Leakey, Richard (1 September 1992). Origins Reconsidered. ISBN 0-385-41264-9.
- Ruhlen, Merritt (1994). The origin of language: tracing the evolution of the mother tongue. New York: Wiley. ISBN 0-471-58426-6.
- Shreeve, James (1995). The Neandertal Enigma: Solving the Mystery of Modern Human Origins. Harper Perennial. ISBN 0-670-86638-5.
- Tattersall, Ian; Schwartz, Jeffrey (2000). Extinct Humans. Boulder and Cumnor Hill: Westview Press. ISBN 0-8133-3482-9.
- Wood, Bernard; Collard, Mark (2001). "The Meaning of Homo". Ludus Vitalis. 9.
- Aiello, Leslie C.; Dunbar, R. I. M. (1993). "Neocortex Size, Group Size, and the Evolution of Language". Current Anthropology. 34 (2): 184–193. doi:10.1086/204160. S2CID 144347664.
- Aiello, Leslie C.; Wells, Jonathan C. K. (2002). "Energetics and the Evolution of the Genus Homo". Annual Review of Anthropology. 31: 323–338. doi:10.1146/annurev.anthro.31.040402.085403.
- Antón, Susan C. (2003). "Natural history of Homo erectus". Yearbook of Physical Anthropology. 46: 126–170. doi:10.1002/ajpa.10399. PMID 14666536.
- Baab, Karen L. (2008). "The taxonomic implications of cranial shape variation in Homo erectus". Journal of Human Evolution. 54 (6): 827–847. doi:10.1016/j.jhevol.2007.11.003. PMID 18191986.
- Bonde, Niels (2012). "Hominid Diversity and 'Ancestor' Myths". In Schilhab, Theresa; Stjernfelt, Frederik; Deacon, Terrence (reds.). The Symbolic Species Evolved. Springer. ISBN 978-94-007-2336-8.
- Ben-Dor, Miki; Gopher, Avi; Hershkovitz, Israel; Barkai, Ran (2011). "Man the Fat Hunter: The Demise of Homo erectus and the Emergence of a New Hominin Lineage in the Middle Pleistocene (ca. 400 kyr) Levant". PLOS ONE. 6 (12): e28689. Bibcode:2011PLoSO...628689B. doi:10.1371/journal.pone.0028689. PMC 3235142. PMID 22174868.
- Brown, Frank; Harris, John; Leakey, Richard; Walker, Alan (1985). "Early Homo erectus skeleton from west Lake Turkana, Kenya". Nature. 316 (6031): 788–792. Bibcode:1985Natur.316..788B. doi:10.1038/316788a0. PMID 3929141. S2CID 4311887.
- Caspari, Rachel; Lee, Sang-Hee (2004). "Older age becomes common late in human evolution". PNAS. 101 (30): 10895–10900. doi:10.1073/pnas.0402857101. PMC 503716. PMID 15252198.
- Ciochon, Russell (2009). "The mystery ape of Pleistocene Asia" (PDF). Nature. 459 (7249): 910–911. Bibcode:2009Natur.459..910C. doi:10.1038/459910a. PMID 19536242. S2CID 205047272.
- Coqueugniot, H.; Hublin, J.-J.; et al. (2004). "Early brain growth in Homo erectus and implications for cognitive ability". Nature. 431 (7006): 299–302. Bibcode:2004Natur.431..299C. doi:10.1038/nature02852. PMID 15372030. S2CID 4428043.
- Dávid-Barrett, Tamás; Dunbar, R. I. M. (2016). "Bipedality and hair loss in human evolution revisited: The impact of altitude and activity scheduling". Journal of Human Evolution. 94: 72–82. doi:10.1016/j.jhevol.2016.02.006. PMC 4874949. PMID 27178459.
- Dennell, Robin; Roebroeks, Wil (2005). "An Asian perspective on early human dispersal from Africa". Nature. 438 (7071): 1099–1104. Bibcode:2005Natur.438.1099D. doi:10.1038/nature04259. PMID 16371999. S2CID 4405913.
- Giles, James (2010). "Naked Love: The Evolution of Human Hairlessness". Biological Theory. 5 (4): 326–336. doi:10.1162/BIOT_a_00062. S2CID 84164968.
- Gowlett, J. A. J. (2016). "The discovery of fire by humans: a long and convoluted process". Philosophical Transactions B. 371 (20150164): 20150164. doi:10.1098/rstb.2015.0164. PMC 4874402. PMID 27216521.
- Hatala, Kevin G.; Roach, Neil T.; Ostrofsky, Kelly R.; Wunderlich, Roshna E.; Dingwall, Heather L.; Villmoare, Brian A.; Green, David J.; Harris, John W. K.; Braun, David R.; Richmond, Brian G. (2016). "Footprints reveal direct evidence of group behavior and locomotion in Homo erectus". Scientific Reports. 6 (28766): 28766. Bibcode:2016NatSR...628766H. doi:10.1038/srep28766. PMC 4941528. PMID 27403790.
- Herries, Andy I. R.; et al. (2020). "Contemporaneity of Australopithecus, Paranthropus, and early Homo erectus in South Africa". Science. 368 (6486): eaaw7293. doi:10.1126/science.aaw7293. hdl:11568/1040368. PMID 32241925. S2CID 214763272.
- Janssen, Marco A.; Sept, Jeanne M.; Griffith, Cameron S. (2007). "Hominids Foraging in a Complex Landscape: Could Homo ergaster and Australopithecus boisei Meet Their Calories Requirements?". In Takahashi, Shingo; Sallach, David; Rouchier, Juliette (reds.). Advancing Social Simulation: The First World Congress. Springer. ISBN 978-4-431-73150-4.
- Kimbel, William H.; White, Tim D. (2017). "Variation, Sexual Dimorphism and the Taxonomy of Australopithecus". In Grine, Frederick E. (red.). Evolutionary History of the Robust Australopithecines. Routledge. ISBN 978-0-202-36137-6.
- Klein, Richard (2005). "Hominin dispersals in the Old World". In Chris, Scarre (red.). The Human Past: World Prehistory & the Development of Human Societies. Thames & Hudson. ISBN 978-0500285312.
- Larick, Roy; Ciochon, Russell (1996). "The African Emergence and Early Asian Dispersals of the Genus Homo". American Scientist. 84 (6): 538–551. Bibcode:1996AmSci..84..538L.
- Latimer, Bruce; Ohman, James C. (2001). "Axial dysplasia in Homo erectus". Journal of Human Evolution. 40: A12.
- Meyer, Marc R.; Vekua, Abesalom; Lordkipanidze, David (2006). "Language and empathy in Homo erectus: behaviors suggested by a modern spinal cord from Dmanisi, but not Nariokotome".
{{cite journal}}: Cite journal requires|journal=(hulp) - Nowell, April; Chang, Melanie Lee (2009). "The Case Against Sexual Selection as an Explanation of Handaxe Morphology" (PDF). PaleoAnthropology. 2009: 77–88.
- Pagel, Mark; Bodmer, Walter (2004). "The Evolution of Human Hairlessness: Cultural Adaptations and the Ectoparasite Hypothesis". In Wasser, Solomon P. (red.). Evolutionary Theory and Processes: Modern Horizons. Springer. doi:10.1007/978-94-017-0443-4_17. ISBN 978-90-481-6457-8.
- Reno, Philip L.; Meindl, Richard S.; McCollum, Melanie A.; Lovejoy, C. Owen (2003). "Sexual dimorphism in Australopithecus afarensis was similar to that of modern humans" (PDF). PNAS. 100 (16): 9404–9409. Bibcode:2003PNAS..100.9404R. doi:10.1073/pnas.1133180100. PMC 170931. PMID 12878734.
- Roberts, Alice (2018). Evolution: The Human Story (Revised uitg.). Dorling Kindersley Ltd. ISBN 978-0-2413-0431-0.
- Rogers, Alan R.; Iltis, David; Wooding, Stephen (2004). "Genetic Variation at the MC1R Locus and the Time since Loss of Human Body Hair". Current Anthropology. 45 (1): 105–108. doi:10.1086/381006. S2CID 224795768.
- Ruxton, Graeme D.; Wilkinson, David M. (2011). "Avoidance of overheating and selection for both hair loss and bipedality in hominins" (PDF). PNAS. 108 (52): 20965–20969. Bibcode:2011PNAS..10820965R. doi:10.1073/pnas.1113915108. PMC 3248486. PMID 22160694.
- Sandgathe, Dennis M.; Berna, Francesco (2017). "Fire and the Genus Homo: An Introduction to Supplement 16". Current Anthropology. 58 (Supplement 16): S165–S175. doi:10.1086/691424. S2CID 164992270.
- Schiess, Regula; Häusler, Martin (2013). "No skeletal dysplasia in the Nariokotome boy KNM-WT 15000 (Homo erectus) – a reassessment of congenital pathologies of the vertebral column". American Journal of Physical Anthropology. 150 (3): 365–374. doi:10.1002/ajpa.22211. PMID 23283736.
- Schiess, Regula; Böni, Thomas; Rühli, Frank J.; Häusler, Martin (2014). "Revisiting scoliosis in the KNM-WT 15000 Homo erectus skeleton" (PDF). Journal of Human Evolution. 67: 48–59. doi:10.1016/j.jhevol.2013.12.009. PMID 24491377.
- Schwartz, Jeffrey H. (2000). "Taxonomy of the Dmanisi Crania" (PDF). Science. 289 (5476): 55–56. doi:10.1126/science.289.5476.55b. PMID 10928927. S2CID 23195350.
- Simpson, Scott W.; Quade, Jay; Levin, Naomi E.; Butler, Robert; Dupont-Nivet, Guillaume; Everett, Melanie; Semaw, Sileshi (2008). "A Female Homo erectus Pelvis from Gona, Ethiopia". Science. 322 (5904): 1089–1092. Bibcode:2008Sci...322.1089S. CiteSeerX 10.1.1.710.7337. doi:10.1126/science.1163592. PMID 19008443. S2CID 22191315.
- Strait, David; Grine, Frederick; Fleagle, John (2015). "Analyzing Hominin Hominin Phylogeny: Cladistic Approach" (PDF). In Henke, Winfried; Tattersall, Ian (reds.). Handbook of Paleoanthropology (2nd uitg.). Springer. doi:10.1007/978-3-642-39979-4_58. ISBN 978-3-642-39979-4. Geargiveer vanaf die oorspronklike (PDF) op 12 Junie 2020. Besoek op 23 Mei 2020.
- Tattersall, Ian (2013). "Homo ergaster and Its Contemporaries" (PDF). In Henke, Winfried; Tattersall, Ian (reds.). Handbook of Paleoanthropology. Springer. ISBN 978-3-642-39978-7. Geargiveer vanaf die oorspronklike (PDF) op 12 Junie 2020. Besoek op 23 Mei 2020.
- Ungar, Peter S.; Grine, Frederick E.; Teaford, Mark F. (2008). "Diet in Early Homo: A Review of the Evidence and a New Model of Adaptive Versatility". Annual Review of Anthropology. 35 (1): 208–228.
- Willems, Erik P.; van Schaik, Carel P. (2017). "The social organization of Homo ergaster: Inferences from anti-predator responses in extant primates". Journal of Human Evolution. 109: 11–21. doi:10.1016/j.jhevol.2017.05.003. PMID 28688456.
- Wang, Weijie; Crompton, Robin H.; Carey, Tanya S.; Günther, Michael M.; Li, Yu; Savage, Russell; Sellers, Williams I. (2004). "Comparison of inverse-dynamics musculo-skeletal models of AL 288-1 Australopithecus afarensis and KNM-WT 15000 Homo ergaster to modern humans, with implications for the evolution of bipedalism". Journal of Human Evolution. 47 (6): 453–478. doi:10.1016/j.jhevol.2004.08.007. PMID 15566947.
- Wood, Bernard; Collard, Mark (1999). "The Human Genus". Science. 284 (5411): 65–71. Bibcode:1999Sci...284...65.. doi:10.1126/science.284.5411.65. PMID 10102822.
- Zhu, Zhaoyu; Dennell, Robin; Huang, Weiwen; Wu, Yi; Qiu, Shifan; Yang, Shixia; Rao, Zhiguo; Hou, Yamei; Xie, Jiubing; Han, Jiangwei; Ouyang, Tingping (2018). "Hominin occupation of the Chinese Loess Plateau since about 2.1 million years ago". Nature. 559 (7715): 608–612. Bibcode:2018Natur.559..608Z. doi:10.1038/s41586-018-0299-4. PMID 29995848. S2CID 49670311.
Skakels
[wysig | wysig bron]- Archaeology Info
- Smithsonian
Wikimedia Commons het meer media in die kategorie Homo ergaster.
Hierdie artikel is in sy geheel of gedeeltelik uit die Engelse Wikipedia vertaal.