Fagocitosi

La fagocitosi ve del grec i significa –fagos, "el que menja" i kytos, "cèl·lula". És un tipus especial d'endocitosi per la qual algunes cèl·lules (neutròfils i macròfags) envolten a un antigen amb la seva membrana citoplasmàtica i l'introdueixen a l'interior de la cèl·lula. Això es produeix gràcies a l'emissió de pseudòpodes al voltant de la partícula o microorganisme fins a envoltar-la completament i formar al voltant d'ell una vesícula, anomenada fagosoma, la qual fusionen posteriorment amb el lisosoma per degradar l'antigen fagocitat.
És un mètode de nutrició, ingestió de matèria de l'exterior (bacteris, altres cèl·lules, matèria inorgànica, etc.), com és el cas d'alguns organismes unicel·lulars. És un dels mitjans de transport gruixut que utilitzen para la seva defensa algunes cèl·lules dels organismes pluricel·lulars. En organismes pluricel·lulars, aquest procés el duen a terme cèl·lules especialitzades, gairebé sempre amb la finalitat de defendre al conjunt de l'organisme davant a potencials invasors perjudicials. En molts organismes superiors, la fagocitosi és tant un mitjà de defensa davant de microorganismes invasors com d'eliminació (fins i tot de reciclatge) de teixits morts.
La fagocitosi fou descoberta el 1882 pel zoòleg rus Il'ja I.Mečnikov, que estudià els processos fagocitaris i veié que en els éssers més simples serveixen especialment per a l'alimentació, en els més complexos faciliten la destrucció d'òrgans i teixits larvals dins els fenòmens de la metamorfosi i, en general, representen una defensa antiinfecciosa important.
Funcions generals de la fagocitosi pels diferents regnes
[modifica]
Els protozous, com és ara les amebes, presenten fagocitosi englobant amb els pseudopodis partícules que empren per a l'alimentació. En els metazous, com ara els porífers, els platihelmints i els celenterats, intervenen fagòcits per ajudar a la digestió dels aliments. La fagocitosi també pot servir per a l'excreció, com ara en els equinoderms i les esponges, on els fagòcits expulsen els catabòlits a l'exterior o hi són expulsats amb ells.
Finalment, la fagocitosi és un dels mecanismes fonamentals de la immunitat natural, especialment en els organismes superiors. En molts indrets del cos (sang, teixit conjuntiu, fetge, etc.) hi ha nombrosos fagòcits, que poden ésser macròfags, com ara els histiòcits i els leucòcits mononuclears, que engloben les partícules grosses, i micròfags, com és ara els leucòcits polinuclears, que són els primers que ataquen qualsevol alteració de l'organisme i formen el pus. En tots els processos inflamatoris els mecanismes fagocitaris, mitjançant micròfags i macròfags, s'encarreguen de destruir o de neutralitzar els cossos estranys (sobretot microbis) que poden posar en perill el funcionament de l'organisme.
Etapes de la fagocitosi
[modifica]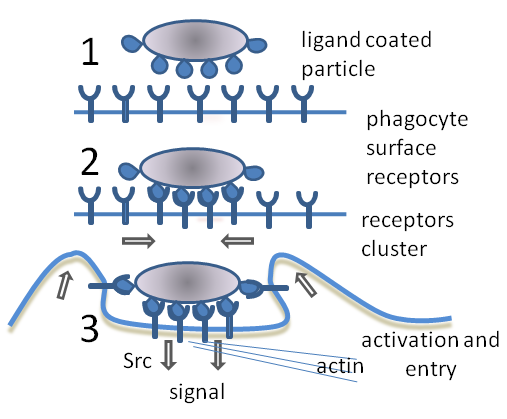
Passatge de cèl·lules al torrent sanguini
[modifica]S'inicia amb l'adhesió de cèl·lules a l'endoteli vascular. Les cèl·lules aniran al lloc de l'amenaça. Aquestes són cèl·lules especialitzades, que poden ser Macròfags o limfòcits. Ells mateixos seran estimulats perquè produeixin citocines (IL-1, TNF, IFN).
Quimiotaxi
[modifica]La quimiotaxi és una etapa de reclutament i mobilització dels leucòcits al teixit o zona lesionada a través d'interaccions cel·lulars. A través d'unions moleculars de baixa afinitat entre receptors en les selectines i els leucòcits sobre la superfície endotelial (selectina E i selectina P, per exemple), el fagòcit s'adhereix lleument a la superfície de l'endoteli (prèviament activat per les citocines). Els leucòcits així adherits, són empesos pel flux sanguini laminar en direcció del corrent sanguini. Mentre que el fagocit es desenganxa de les interaccions corrent-amunt, els seus lligands de membrana s'uneixen a corrent-avall a noves selectines. El resultat és un moviment net a través de la superfície endotelial. Les molècules d'adhesió vascular (VCAM-1) presents a l'endoteli són unes altres molècules que participen en aquesta mobilització, que els seus lligands corresponents mostren preferència pels limfòcits T i eosinòfils. Arribarà un moment que, amb la presència i l'activació de quimiocines, els fagòcits mobilitzants estableixen interaccions intercel·lulars de gran afinitat amb l'endoteli per mitjà d'integrines i altres lligands endotelials. En especial les molècules endotelials LFA-a, CR3 i VLA-4 s'adhereixen a lligands específics sobre els fagocits, entre ells VCAM-1 i ICAM-1. Les proteïnes inflamatòries com el TNF i la IL-1, són les encarregades de regular l'expressió d'aquests lligands sobre la superfície del fagòcit. A partir d'aquí, els fàgocits, atrets pel gradient de concentració de les quimiocines, travessen l'epiteli vascular cap al focus d'infecció patògena.
Adherència
[modifica]Trobarem que molts altres fagòcits i receptors de sobre la membrana dels leucòcits actuen com a mecanisme d'adhesió sobre els microorganismes, sigui sobre opsonines del sistema immunitari de l'hoste com productes microbians específics.
- Receptors de mannosa: El receptor de la mannosa té molta afinitat pels components que contenen mannosa presents en les glicoproteïnes i glicolípids de les parets cel·lulars microbianes.
- També trobem els receptors Scavenger que s'uneixen directament a molècules de LDL modificades i a microorganismes.
- Trobarem el CD14 que és un lligand amb preferència específica als lipopolisacàrids presents en certs bacteris i està associat a un receptor de tipus Toll.
- Un receptor recentment descobert és la transmembrana de set hèlix d'alfa que, la seva funció principal està associada a senyals de certs pèptids microbians i a senyals de quimiocines.
- Hi ha receptors pels fragments Fc dels anticossos opsonizans IgG2 i IgG3.
Ingestió
[modifica]Sabem que, la unió a receptors d'adherència promou senyals de comunicació intracel·lular que sorgeixen en la invaginació de la membrana del fagòcit rodejant al receptor i el seu lligand patogènic. Al rodejar per complet el complex receptor-molècula, la membrana s'uneix als seus extrems i allibera a l'interior de la cèl·lula un fagosoma. Això pot passar en més d'un punt de la membrana cel·lular.
Digestió
[modifica]En el citoplasma, quan ja ha arribat el fagosoma, es produeix la seva desintegració, que es pot realitzar a través de diversos mecanismes dependents o independents d'oxigen. El primer mecanisme, el d'oxigen, es dona després que rutes que consumeixen oxigen s'hagin activat, que produeix l'alliberació de radicals lliures de l'oxigen que són tòxics pels microorganismes. En el segon cas, l'independent d'oxigen, és on intervenen els lisosomes, els quals s'uneixen al fagosoma formant un fagolisosoma i alliberant enzims hidrolítics que destruiran l'antigen.
Excreció
[modifica]Durant el procés de digestió, trobem una vesícula que conté desfets o antígens (ja que, aquest no sempre pot ser desintegrat), que ha d'estar fora de la cèl·lula per portar futurs inconvenients. Així que la millor forma per eliminar o desfer-se d'aquests residus és a través de l'exocitosi. Per entendre millor aquest terme es pot posar un exemple molt clar: quan nosaltres tossim el que estem fent és desfer-nos de cèl·lules que contenen un antigen que no podem degradar. Aquestes cèl·lules anteriors són els macròfags alveolars, que quan entra una partícula exògena que no poden degradar es tornen una amenaça pel nostre organisme, per això, el que més ens convé és desfer-nos d'aquestes cèl·lules.
Autofagosoma
[modifica]Totes les cèl·lules eucariotes posseeixen autofagosomes, sobretot aquelles cèl·lules que estan danyades i desnodrides.
Els autofagosomes estan formats per una membrana doble. Aquests parteixen d'una organització citoplasmàtica de lípids i proteïnes anomenat també fagòfor, que creix i pren una forma còncava. La formació d'autofagosomes és iniciada per la classe III de fosfoinositol-3-kinasa i per un gen relacionat amb l'autofàgia (Atg) 6 (també conegut com a Beclin-1). A més a més, dos sistemes més llunyans estan implicats, per una banda la proteïna tipus ubiquitina Atg8 (conegut com a LC3 en cèl·lules dels mamífers) i la proteasa Atg4 i, per una altra banda el complex de Atg12-Atg5-Atg16.
Els autofagosomes, primer, engloben components cel·lulars que s'han de destruir per després tancar-se en forma de vesícules de doble membrana amb una mida variable. Aquests poden reciclar el contingut del seu interior que creguin útil. El fagosoma té les següents funcions:
- Neteja i reciclatge cel·lular: Digestió intracel·lular d'orgànuls danyats.
- Obtenció de material en períodes de manca de nutrients. Autodigestió i disminució de la mida.
- Duen a terme l'eliminació cel·lular quan no funciona l'apoptosi.
- Elimina els patògens intracel·lulars.
- Són desencadenants de la resposta immunitària innata davant de paràsits intracel·lulars.
- Presentació d'antígens interns pel MHC II.
No coneixem gaire la seva biogènesi, es pensa que pot ésser per autosemblatge de components del citoplasma o per la unió de membranes procedents d'altres orgànuls, com la de l'Aparell de Golgi o la del Reticle endoplasmàtic. Però coneixem que el fagosoma es pot unir amb el lisosoma i formar un autofagolisosoma (el mateix que passa amb el fagosoma) que el seu contingut es pot degradar mitjançant hidrolases àcides lisosomals.
Referències
[modifica]- Enciclopèdia lliure universal
- Gran Enciclopèdia Catalana
- Vídeo de la fagocitosi Arxivat 2014-06-26 a Wayback Machine.
- Bioquímica: fundamentos para medicina y ciencias de la vida; Werner Müller-Esterl, ed. Reverté
- Medline plus «MedlinePlus - Resultados sobre: fagocitosis».
| Orgànuls i estructures cel·lulars | Acrosoma · Aparell de Golgi · Centríol · Cili · Citoplasma · Cloroplast · Citoesquelet · Endosoma · Flagel · Glioxisoma · Leucoplast · Lisosoma · Melanosoma · Membrana nuclear · Membrana plasmàtica · Mitocondri · Nuclèol · Nucli cel·lular · Peroxisoma · Reticle endoplasmàtic · Ribosoma · Vacúol · Vesícula · VTC's |
|---|---|
| Processos cel·lulars | |
| Metabolisme de les macromolècules | |