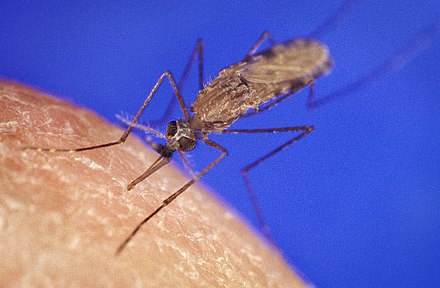
Anopheles gambiae
| Anopheles gambiae | |
|---|---|
| |
| Scientific classification | |
| Kingdom: | |
| Phylum: | |
| Class: | |
| Order: | |
| Family: | |
| Subfamily: | |
| Genus: | |
| Species complex: | A. gambiae sensu lato
|
| Species: |
|
| Binomial name | |
| Anopheles gambiae | |

The Anopheles gambiae complex consists of at least seven morphologically indistinguishable species of mosquitoes in the genus Anopheles. The complex was recognised in the 1960s and includes the most important vectors of malaria in sub-Saharan Africa, particularly of the most dangerous malaria parasite, Plasmodium falciparum.[2] It is one of the most efficient malaria vectors known. The An. gambiae mosquito additionally transmits Wuchereria bancrofti which causes lymphatic filariasis, a symptom of which is elephantiasis.[3]
Discovery and elements
[edit]The Anopheles gambiae complex or Anopheles gambiae sensu lato was recognized as a species complex only in the 1960s. The A. gambiae complex consists of:
- Anopheles arabiensis
- Anopheles bwambae
- Anopheles melas
- Anopheles merus
- Anopheles quadriannulatus[4][5]
- Anopheles gambiae sensu stricto (s.s.)[6]
- Anopheles coluzzii
- Anopheles amharicus
The individual species of the complex are morphologically difficult to distinguish from each other, although it is possible for larvae and adult females. The species exhibit different behavioural traits. For example, Anopheles quadriannulatus is both a saltwater and mineralwater species. A. melas and A. merus are saltwater species, while the remainder are freshwater species.[7] Anopheles quadriannulatus generally takes its blood meal from animals (zoophilic), whereas Anopheles gambiae sensu stricto generally feeds on humans, i.e. is considered anthropophilic.[citation needed] Identification to the individual species level using the molecular methods of Scott et al. (1993)[8] can have important implications in subsequent control measures.
Anopheles gambiae in the strict sense
[edit]An. gambiae sensu stricto (s.s.) has been discovered to be currently in a state of diverging into two different species—the Mopti (M) and Savannah (S) strains—though as of 2007, the two strains are still considered to be a single species.[9][10]
A mechanism of species recognition using the sound emitted by the wings and identified by Johnston's organ was proposed in 2010,[11] however this mechanism has never been confirmed since, and the overall mechanism theory through "harmonic convergence" has been challenged.[12][13]
Genome
[edit]An. gambiae s.s. genomes have been sequenced three times, once for the M strain, once for the S strain, and once for a hybrid strain.[9][10] Currently, ~90 miRNA have been predicted in the literature (38 miRNA officially listed in miRBase) for An. gambiae s.s. based upon conserved sequences to miRNA found in Drosophila.[citation needed] Holt et al., 2002 and Neafsey et al., 2016 find transposable elements to be ~13% of the genome, similar to Drosophila melanogaster (also in Diptera).[14] However they find the proportion of TE types to be very different from D. melanogaster with approximately the same composition of long terminal repeat retrotransposons, non-long terminal repeat retrotransposons and DNA transposons.[14] These proportions are believed to be representative of the genus.[14]
The genetics and genomics of sex chromosomes have been discovered and studied by Windbichler et al., 2007 and Galizi et al., 2014 (a Physarum polycephalum homing endonuclease which destroys X chromosomes), Windbichler et al., 2008 and Hammond et al., 2016 (methods to reduce the female population), Windbichler et al., 2011 (trans from yeast), Bernardini et al., 2014 (a method to increase the male population), Kyrou et al., 2018 (a female necessary exon and a homing endonuclease to drive it), Taxiarchi et al., 2019 (sex chromosome dynamics in general) and Simoni et al., 2020 (an X chromosome destroying site specific nuclease).[15] See § Gene drive below for their applications.
An. gambiae has a high degree of polymorphism. This is especially true in the cytochrome P450s, Wilding et al., 2009 finding 1 single nucleotide polymorphism (SNP)/26 base pairs. This species has the highest amount of polymorphism in the CYPs of any insect known, much tending to be found in "scaffolds" that are found only in particular subpopulations. These are termed "dual haplotype regions" by Holt et al., 2002 who sequenced the PEST strain.[16]: 241
In common with many chromosomes, An. gambiae codes for spindle and kinetochore-associated proteins. Hanisch et al., 2006 locate AgSka1, the spindle and kinetochore-associated protein 1 gene, at EAL39257.[17]
The entire Culicidae family may or may not conserve epigenetic mechanisms – as of 2012[update] this remains unresolved.[18] Toward answering this question, Marhold et al., 2004 compare their own previous work in Drosophila melanogaster against new sequences of D. pseudoobscura and An. gambiae.[18] They find all three do share the DNA methylation enzyme DNMT2 (DmDNMT2, DpDNMT2, and AgDNMT2).[18] This suggests all Diptera may conserve an epigenetic system employing Dnmt2.[18]
Hosts
[edit]Hosts include Bos taurus, Capra hircus, Ovis aries and Sus scrofa.[19]
Parasites
[edit]Parasites include Plasmodium berghei (for which it also serves as a vector),[20][21][22] and the bioinsecticides/entomopathogenic fungi Metarhizium robertsii[20] and Beauveria bassiana.[20] All three of these parasites combine with insecticides to reduce fitness – see § Insecticides below.[20] CRISPR/Cas9 and U6-gRNA are increasingly (as of 2020[update]) being used together for knockout experiments in mosquitoes.[22] Dong et al., 2018 develops and presents a new U6-gRNA+Cas9 technique in An. gambiae, and utilizes it to knock out fibrinogen related protein 1 (FREP1), thereby severely reducing infection of the mosquito by P. berghei and P. falciparum.[22] However this also demonstrates the centrality of FREP1 to the insect's success, impairing all measured activities across all life stages.[22] Yang et al., 2020 uses the Dong method to do the same with mosGILT, also severely reducing Plasmodium infection of the mosquito but also finding a vital life process is impaired, in mosGILT's case ovary development.[22]
Control
[edit]Insecticides
[edit]Parasites/bioinsecticides and chemical insecticides synergistically reduce fitness. Saddler et al., 2015 finds even An. gambiae with knockdown resistance (kdr) are more susceptible to DDT if they are first infected with Plasmodium berghei[20][21] and Farenhorst et al., 2009 the same for Metarhizium robertsii or Beauveria bassiana.[20] This is probably due to an effect found by Félix et al., 2010 and Stevenson et al., 2011: An. gambiae alters various activities – especially CYP6M2 – in response to P. berghei invasion. CYP6M2 is known to somehow produce pyrethroid resistance, and pyrethroids and DDT share a mechanism of action.[21]
Gene drive
[edit]Research relevant to the development of gene drive controls of An. gambiae have been performed by Windbichler et al., 2007, Windbichler et al., 2008, Windbichler et al., 2011, Bernardini et al., 2014, Galizi et al., 2014, Hammond et al., 2016, Kyrou et al., 2018, Taxiarchi et al., 2019 and Simoni et al., 2020.[15] For specific genes involved see § Genome above. These can all be used in pest control because they induce infertility.[15]
Fecundity
[edit]Fecundity of An. gambiae depends on the detoxification of reactive oxygen species (ROS) by catalase.[23] Reduction in catalase activity significantly reduces reproductive output of female mosquitoes, indicating that catalase plays a central role in protecting oocytes and early embryos from ROS damage.[23]
Historical note
[edit]An. gambiae invaded northeastern Brazil in 1930, which led to a malaria epidemic in 1938/1939.[24] The Brazilian government assisted by the Rockefeller Foundation in a programme spearheaded by Fred Soper eradicated these mosquitoes from this area. This effort was modeled on the earlier success in eradication of Aedes aegypti as part of the yellow fever control program. The exact species involved in this epidemic has been identified as An. arabiensis.[25]
Peptide hormones
[edit]Kaufmann and Brown 2008 find the An. gambiae adipokinetic hormone (AKH) mobilizes carbohydrates but not lipids. Meanwhile AKH/Corazonin Peptide (ACP) does not mobilize (or inhibit mobilization) of either. Mugumbate et al., 2013 provides in solution and membrane bound structures from a nuclear magnetic resonance investigation.[26]
References
[edit]- ^ Giles, G. M. (1902). A handbook of the gnats or mosquitoes giving the anatomy and life history of the Culicidae together with descriptions of all species noticed up to the present date. London, United Kingdom: John Bale, Sons & Danielsson.
- ^ "Anopheles gambiae complex". Walter Reed Army Institute of Research. Archived from the original on 2007-09-29.
- ^ "Lymphatic filariasis". www.who.int. Retrieved 2020-04-10.
- ^ Besansky NJ, Powell JR, Caccone A, Hamm DM, Scott JA, Collins FH (July 1994). "Molecular phylogeny of the Anopheles gambiae complex suggests genetic introgression between principal malaria vectors". Proceedings of the National Academy of Sciences. 91 (15): 6885–8. Bibcode:1994PNAS...91.6885B. doi:10.1073/pnas.91.15.6885. PMC 44302. PMID 8041714.
- ^ Wilkins EE, Howell PI, Benedict MQ (2006). "IMP PCR primers detect single nucleotide polymorphisms for Anopheles gambiae species identification, Mopti and Savanna rDNA types, and resistance to dieldrin in Anopheles arabiensis". Malaria Journal. 5 (1): 125. doi:10.1186/1475-2875-5-125. PMC 1769388. PMID 17177993.
- ^ Yakob, Laith (2011). "Epidemiological consequences of a newly discovered cryptic subgroup of Anopheles gambiae". Biology Letters. 7 (6): 947–949. doi:10.1098/rsbl.2011.0453. PMC 3210673. PMID 21693489.
- ^ G.B. White (1974). "Anopheles gambiae complex and disease transmission in Africa". Transactions of the Royal Society of Tropical Medicine and Hygiene. 68 (4): 278–298. doi:10.1016/0035-9203(74)90035-2. PMID 4420769.
- ^ C. Fanello; F. Santolamazza; A. Della Torre (2002). "Simultaneous identification of species and molecular forms of the Anopheles gambiae complex by PCR-RFLP". Medical and Veterinary Entomology. 16 (4): 461–4. doi:10.1046/j.1365-2915.2002.00393.x. PMID 12510902. S2CID 28983355.
- ^ a b "Anopheles gambiae: First genome of a vector for a parasitic disease". Genoscope. Archived from the original on 2011-08-07.
- ^ a b Lawniczak, M. K.; et al. (Oct 22, 2010). "Widespread divergence between incipient Anopheles gambiae species revealed by whole genome sequences". Science. 330 (6003): 512–4. Bibcode:2010Sci...330..512L. doi:10.1126/science.1195755. PMC 3674514. PMID 20966253.
- ^ Pennetier, Cédric; Warren, Ben; Dabiré, K. Roch; Russell, Ian J.; Gibson, Gabriella (2010). ""Singing on the Wing" as a Mechanism for Species Recognition in the Malarial Mosquito Anopheles gambiae". Current Biology. 20 (2): 131–136. Bibcode:2010CBio...20..131P. doi:10.1016/j.cub.2009.11.040. PMID 20045329. S2CID 15185976.
- ^ Somers, Jason; Georgiades, Marcos; Su, Matthew P.; Bagi, Judit; Andrés, Marta; Alampounti, Alexandros; Mills, Gordon; Ntabaliba, Watson; Moore, Sarah J.; Spaccapelo, Roberta; Albert, Joerg T. (2022-01-14). "Hitting the right note at the right time: Circadian control of audibility in Anopheles mosquito mating swarms is mediated by flight tones". Science Advances. 8 (2): eabl4844. Bibcode:2022SciA....8.4844S. doi:10.1126/sciadv.abl4844. ISSN 2375-2548. PMC 8754303. PMID 35020428.
- ^ Feugère, L.; Simões, P.M.V.; Russell, I.J.; Gibson, G. (2022-12-18), Ignell, R.; Lazzari, C.R.; Lorenzo, M.G.; Hill, S.R. (eds.), "Chapter 26: The role of hearing in mosquito behaviour", Sensory ecology of disease vectors, Brill | Wageningen Academic, pp. 683–708, doi:10.3920/978-90-8686-932-9_26, ISBN 978-90-8686-380-8, retrieved 2024-05-30
- ^ a b c Gilbert, Clément; Peccoud, Jean; Cordaux, Richard (2021-01-07). "Transposable Elements and the Evolution of Insects" (PDF). Annual Review of Entomology. 66 (1): 355–372. doi:10.1146/annurev-ento-070720-074650. ISSN 0066-4170. PMID 32931312. S2CID 221747772.
- ^ a b c Hay, Bruce A.; Oberhofer, Georg; Guo, Ming (2021-01-07). "Engineering the Composition and Fate of Wild Populations with Gene Drive". Annual Review of Entomology. 66 (1): 407–434. doi:10.1146/annurev-ento-020117-043154. ISSN 0066-4170. PMID 33035437. S2CID 222257628.
- ^ Gilbert, Lawrence I., ed. (2012). Insect molecular biology and biochemistry. Amsterdam Boston: Academic Press. pp. x+563. ISBN 978-0-12-384747-8. OCLC 742299021.
- ^ Cheeseman, Iain M.; Desai, Arshad (2008). "Molecular architecture of the kinetochore–microtubule interface". Nature Reviews Molecular Cell Biology. 9 (1). Nature Portfolio: 33–46. doi:10.1038/nrm2310. ISSN 1471-0072. PMID 18097444. S2CID 34121605.
- ^ a b c d Severson, David W.; Behura, Susanta K. (2012-01-07). "Mosquito Genomics: Progress and Challenges". Annual Review of Entomology. 57 (1). Annual Reviews: 143–166. doi:10.1146/annurev-ento-120710-100651. ISSN 0066-4170. PMID 21942845.
- ^ "Anopheles gambiae". Invasive Species Compendium. CABI. 2019-11-24. Retrieved 2022-01-26.
- ^ a b c d e f Alout, Haoues; Labbé, Pierrick; Chandre, Fabrice; Cohuet, Anna (2017). "Malaria Vector Control Still Matters despite Insecticide Resistance". Trends in Parasitology. 33 (8): 610–618. doi:10.1016/j.pt.2017.04.006. ISSN 1471-4922. PMID 28499699. S2CID 32524464.
- ^ a b c Minetti, Corrado; Ingham, Victoria A; Ranson, Hilary (2020). "Effects of insecticide resistance and exposure on Plasmodium development in Anopheles mosquitoes" (PDF). Current Opinion in Insect Science. 39: 42–49. Bibcode:2020COIS...39...42M. doi:10.1016/j.cois.2019.12.001. ISSN 2214-5745. PMID 32109860. S2CID 211563675.
- ^ a b c d e Caragata, E.P.; Dong, S.; Dong, Y.; Simões, M.L.; Tikhe, C.V.; Dimopoulos, G. (2020-09-08). "Prospects and Pitfalls: Next-Generation Tools to Control Mosquito-Transmitted Disease". Annual Review of Microbiology. 74 (1). Annual Reviews: 455–475. doi:10.1146/annurev-micro-011320-025557. ISSN 0066-4227. PMID 32905752. S2CID 221625690.
- ^ a b DeJong RJ, Miller LM, Molina-Cruz A, Gupta L, Kumar S, Barillas-Mury C (February 2007). "Reactive oxygen species detoxification by catalase is a major determinant of fecundity in the mosquito Anopheles gambiae". Proceedings of the National Academy of Sciences. 104 (7): 2121–6. Bibcode:2007PNAS..104.2121D. doi:10.1073/pnas.0608407104. PMC 1892935. PMID 17284604.
- ^ Killeen GF (October 2003). "Following in Soper's footsteps: northeast Brazil 63 years after eradication of Anopheles gambiae". The Lancet Infectious Diseases. 3 (10): 663–6. doi:10.1016/S1473-3099(03)00776-X. PMID 14522266.
- ^ Parmakelis A, Russello MA, Caccone A, et al. (January 2008). "Historical analysis of a near disaster: Anopheles gambiae in Brazil". The American Journal of Tropical Medicine and Hygiene. 78 (1): 176–8. doi:10.4269/ajtmh.2008.78.176. PMID 18187802.
- ^ Strand, M.R.; Brown, M.R.; Vogel, K.J. (2016). "Mosquito Peptide Hormones". Advances in Insect Physiology. Vol. 51. Elsevier. pp. 145–188. doi:10.1016/bs.aiip.2016.05.003. ISBN 9780128024577. ISSN 0065-2806. PMC 6338476. PMID 30662099. S2CID 58546659.
External links
[edit]- "Anopheles gambiae". VectorBase. Archived from the original on 2019-01-27. Retrieved 2013-07-12.
- View the anoGam1 genome assembly in the UCSC Genome Browser
- DiArk
| Authority control databases: National |
|---|