Banaanikärpänen
| Banaanikärpänen | |
|---|---|
 Naaras |
|
| Tieteellinen luokittelu | |
| Domeeni: | Aitotumaiset Eucarya |
| Kunta: | Eläinkunta Animalia |
| Pääjakso: | Niveljalkaiset Arthropoda |
| Alajakso: | Kuusijalkaiset Hexapoda |
| Luokka: | Hyönteiset Insecta |
| Alaluokka: | Siipikantaiset Pterygota |
| Osaluokka: | Neoptera |
| Lahko: | Kaksisiipiset Diptera |
| Alalahko: | Kärpäset Brachycera |
| Osalahko: | Muscomorpha |
| Heimo: | Mahlakärpäset Drosophilidae |
| Alaheimo: | Drosophilinae |
| Suku: | Drosophila |
| Laji: | melanogaster |
| Kaksiosainen nimi | |
| Synonyymit | |
|
|
| Katso myös | |
Banaanikärpänen (Drosophila melanogaster) on pieni kellanruskea mahlakärpäslaji, joka on jo vuosikymmenten ajan ollut yksi kokeellisen biologisen tutkimuksen tärkeimmistä malliorganismeista. Yleiskielessä banaanikärpäsellä voidaan tarkoittaa myös koko mahlakärpästen heimoa tai useita muita Drosophila-suvun lajeja, joista monet ovat Suomessa huomattavasti D. melanogaster -lajia yleisempiä.
Vaikka banaanikärpänen on tavallinen vieras suomalaisissakin kodeissa, se ei pysty talvehtimaan Suomessa vaan kanta on riippuvainen Baltian maista saapuvista ja kauppojen hedelmien mukana tulevista yksilöistä. Ravinnokseen mahlakärpäslajit imevät käymistilassa olevia nesteitä, esimerkiksi hedelmien mehua ja mahlaa. Toukat elävät esimerkiksi mahlassa tai maahan pudonneissa marjoissa ja hedelmissä syöden nesteitä sekä hiivoja ja muita mikro-organismeja.
Banaanikärpäsen biologisella tutkimuksella on pitkä historia. Nykyäänkin se on yksi yleisimmin käytetyistä malliorganismeista, josta on eristetty laboratoriokasvatuksiin tuhansia genotyypiltään erilaisia mutanttikantoja. Lajin yksilöitä käytetäänkin lähes kaikilla biologian osa-alueilla. Erityisesti banaanikärpäsiä käytetään geneettisissä, kehitysbiologisissa ja fysiologisissa tutkimuksissa, joissa voidaan selvittää muun muassa muistin toimintaa tai alkoholin vaikutuksia. Lajin koko genomin emäsjärjestys selvitettiin jo vuonna 2000. Koska ihmisen ja banaanikärpäsen geenitoimintojen on havaittu olevan pohjimmiltaan melko samanlaisia, voidaan kärpäsellä saatuja tuloksia soveltaa usein myös ihmisellä.
Banaanikärpäsen suosioon malliorganismina ovat vaikuttaneet monet seikat, mutta erityisesti sen helppo kasvatettavuus, taudinkestävyys, lyhyt sukupolvenkierto ja mutaatioiden synnyttämisen helppous nostivat sen tutkijoiden suosioon jo 1900-luvun alussa. Nykyään samoja ominaisuuksia käyttävät hyväkseen muun muassa monet matelija- ja akvaarioharrastajat, jotka käyttävät siivettömiä banaanikärpäsen mutanttikantoja lemmikkiensä elävänä ravintona.
Ulkonäkö
[muokkaa | muokkaa wikitekstiä]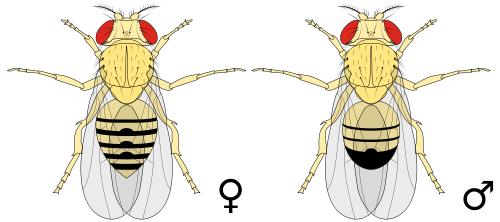
Banaanikärpänen on kellanruskea, raidallinen ja punasilmäinen pieni kärpänen. Banaanikärpänen on sukupuolidimorfinen laji, eli sen koiraan ja naaraan ulkonäöt poikkeavat toisistaan. Naaraat ovat noin 2,5 millimetriä pitkiä, ja koiraat ovat hieman tätä pienempiä. Sukupuolten välisiä muita eroja ovat esimerkiksi koiraan peräaukon ja sukupuolielimien ympärillä olevat piikkimäiset karvat ja koiraan ensimmäisessä raajaparissa sijaitsevat sukaskimput (engl. sex combs), jotka puuttuvat naaraalta. Koiraan takaruumis on kapeampi, sen kärki tummempi eivätkä takaruumiin alapuoliset sukupuolielimet ole yhtä selvästi näkyvillä koirailla kuin naarailla.[3]
Mutanttikannat
[muokkaa | muokkaa wikitekstiä]Banaanikärpäsestä on eristetty laboratoriossa tuhansia erilaisia mutanttikantoja, joiden ulkonäkö saattaa poiketa huomattavasti villityypistä. Esimerkiksi white (w) -mutantin silmät ovat valkoiset ja forked (f) -mutantin selkäpuolen karvat ovat koukkupäisiä. Mutaatioiden seuraukset voivat olla myös huomattavasti dramaattisempia. Esimerkiksi wingless (wg) -mutantit ovat kokonaan siivettömiä.[4] Listaa kaikista tunnetuista ja tutkimuslaitoksissa ylläpidetyistä Drosophilan mutanttikannoista ylläpidetään FlyBase:ssa, jonka kautta tutkijat voivat myös ottaa yhteyttä toisiin laboratorioihin tai kantakeskuksiin tilatakseen itselleen erilaisten mutanttikantojen yksilöitä.
Elinkierto
[muokkaa | muokkaa wikitekstiä]

Banaanikärpäsen elinkierron pituus vaihtelee lämpötilan mukaan kuten kaikilla vaihtolämpöisillä eliöillä. Elinkierto on lyhimmillään 28 °C:ssa, jolloin kärpäsen kehitys munasta aikuiseksi vie seitsemän päivää. Tätä korkeammissa lämpötiloissa kehitys hidastuu korkeamman lämpötilan aiheuttaman fysiologisen stressin vuoksi; esimerkiksi 30 °C:ssa kehitys kestää 11 päivää. Ihanteellisissa olosuhteissa, 25 °C:ssa, kehitys vie 8,5 päivää. Tätä alemmissa lämpötiloissa kehitys hidastuu: 18 °C:ssa kuluu 19 päivää ja 12 °C:ssa kuluu yli 50 päivää.[3] Jos kärpäsiä on samassa paikassa paljon, hidastaa se kärpästen kehitystä.[5] Ne myös jäävät kooltaan pienemmiksi.[6]
Naaraat laskevat suurin piirtein 400 munaa sopivalle alustalle noin viisi kappaletta kerrallaan. Kooltaan 0,5 mm:n mittaiset munat kuoriutuvat 12–15 tunnin kuluttua (25 °C:ssa) ensimmäisen toukkavaiheen toukiksi. Toukat kasvavat noin neljä päivää, luoden nahkansa kaksi kertaa noin 24 ja 48 tunnin kuluttua kuoriutumisesta. Ensimmäisen nahanluonnin jälkeen toukkia kutsutaan toisen toukkavaiheen toukiksi, ja kolmas toukkavaihe alkaa toisen nahanluonnin jälkeen. Tämän jälkeinen kotelovaihe ja metamorfoosi kestävät neljä päivää, jonka kuluttua aikuiset kuoriutuvat. Naaraat ovat parittelukypsiä noin 8–12 tunnin kuluttua kuoriutumisesta. Aikuinen banaanikärpänen elää noin 30 päivää.[3]
Yksilönkehitys ja alkionkehitys
[muokkaa | muokkaa wikitekstiä]Banaanikärpäsen alkionkehitystä eli embryogeneesiä on tutkittu erittäin paljon, ja varhaisen alkion kehitystä säätelevä geeniverkosto on yksi parhaiten tunnetuista säätelyverkostoista. Suosion taustalla ovat olleet samat ominaisuudet, jotka ovat tehneet banaanikärpäsestä genetiikan malliorganismin. Banaanikärpäsen alkionkehitys eroaa tosin muiden malliorganismien kehityksestä. Sen vakoutuminen nimittäin tapahtuu solusitkoksessa, joka on useissa tumanjakautumisissa ilman solunjakautumista syntynyt monitumainen solu. Oogeneesin aikana rengaskanaviksi (engl. ring canals) kutsutut solulimasillat yhdistävät muodostuvan oosyytin syöttösoluihin, joista kulkeutuu oosyyttiin ravinteita ja kehitystä sääteleviä molekyylejä.[7]

Oosyytistä kehittyvän munasolun hedelmöityksen jälkeen varhainen alkio käy läpi useita DNA:n monistumiskertoja, mutta vain muutamia tumanjakautumisia. Vasta myöhemmissä kehityksen vaiheissa alkion solut eroavat erillisiksi soluiksi. Koska DNA:n monistuminen eli replikaatio varhaisessa alkiossa on niin nopeaa, ei solusyklissä ole juurikaan tarkastuspisteitä (engl. checkpoints) estämässä virheellisten solujen muodostumista. Sen vuoksi syntyneet virheelliset solut vaeltavat alkion ruskuaispussiin, joka ei muodosta aikuisen kärpäsen rakenteita. Alkio käy kehityksensä aikana läpi useita vaiheita, jotka ovat erittäin hyvin tunnettuja, kunnes se lopulta kuoriutuu ensimmäisen toukkavaiheen toukkana.[7]
Toukan kehityksen aikana imaginaalilevyinä (engl. imaginal discs) tunnetut kudokset kasvavat toukan sisällä. Imaginaalilevyt muodostavat kehittyessään suurimman osan aikuisen kärpäsen rakenteista, kuten pään, jalat, siivet ja sukupuolielimet. Niiden solut erkanevat muista solulinjoista jo alkionkehityksen aikana ja niiden jakautuminen jatkuu myös toukkavaiheiden aikana, toisin kuin useimpien muiden toukan solukkojen. Kotelovaiheen aikana imaginaalilevyjen kudokset läpikäyvät monia voimakkaita muutoksia, jotta ne voivat muodostaa aikuisen kärpäsen rakenteita.[8]
Elintavat
[muokkaa | muokkaa wikitekstiä]Elinympäristö ja ravinto
[muokkaa | muokkaa wikitekstiä]Banaanikärpänen on kosmopoliitti laji, joka on levinnyt kaikkialle maailmaan.[9][10] Sitä tavataan luonnossa myös Suomessa.[11][9] Laji ei kuitenkaan pysty talvehtimaan Suomessa muutamien muiden suvun lajien tavoin, vaan kanta on riippuvainen Baltian maista saapuvista ja osin myös kauppojen hedelmien mukana tulevista yksilöistä.[9][12] Sen sijaan esimerkiksi lähisukuinen D. littoralis pystyy selviämään talven yli jopa pohjoisimmassa Suomessa.[12] Yhteensä luonnonvaraisia Drosophila-kärpäsiä Suomessa tavataan noin 30 lajia.[9] D. melanogasterin luonnonkanta on runsaimmillaan loppukesästä, jolloin kärpäsiä etsiytyy ihmiskoteihin. Banaanikärpäset, kuten muutkin mahlakärpäset, viihtyvät kosteissa ja varjoisissa paikoissa. Ne ovat aktiivisimmin liikkeellä pääasiassa illalla.[13]
Ravinnokseen mahlakärpäslajit imevät käymistilassa olevia nesteitä, esimerkiksi sokeripitoisia hedelmiä ja puiden mahlaa. Toukat elävät mahlassa, sienissä, maahan pudonneissa marjoissa ja pilaantuneissa hedelmissä syöden niiden nesteitä sekä erityisesti niissä kasvavia hiivoja ja muita mikro-organismeja.[13] Banaanikärpänen pystyy tunnistamaan makean ja kitkerän maun, mutta ei esimerkiksi chilipippurille tyypillistä polttavaa kapsaisiinia.[14]
Kosiokäyttäytyminen ja parittelu
[muokkaa | muokkaa wikitekstiä]Banaanikärpäset pyrkivät parittelemaan mahdollisimman pian kuoriutumisensa jälkeen. Naaraat ovat halukkaita paritteluun jo 8–12 tuntia kuoriutumisesta.[15] Sitä nuorempia, parittelemattomia naaraita kutsutaan usein virgiineiksi naaraiksi.[16] Banaanikärpäskoiraan kosiokäyttäytyminen muodostuu viiden toiminnon sarjasta, jonka se suorittaa houkutellakseen naaraita parittelemaan kanssaan. Ensin koiraat suoristavat itsensä ja värisyttävät siipiään kosioäänen muodostamiseksi. Pian sen jälkeen koiras siirtyy matalana naaraan takaruumiin taakse ja alkaa kosketella naaraan genitaaleja jaloillaan ja imukärsällään. Lopulta koiras käyristää takaruumistaan ja yrittää parittelua. Naaras voi torjua koiraan joko siirtymällä kauemmas tai työntämällä munanasettimensa ulos. Onnistunut parittelu kestää 10 minuuttia, jonka aikana koiras siirtää satoja siittiöitä naaraaseen. Naaras pystyy säilömään spermaa, joka voi joutua siten myös kilpailemaan munasolujen hedelmöityksestä muiden koiraiden siittiöiden kanssa.[17]
Lento
[muokkaa | muokkaa wikitekstiä]Banaanikärpänen on useimpien kärpäslajien tavoin erittäin taitava lentäjä. Sen siivet pystyvät lyömään jopa 220 kertaa sekunnissa.[18] Lentävä banaanikärpänen on myös hyvin ketterä, sillä se voi muuttaa lentosuuntaansa yhdeksänkymmentä astetta jopa alle viidessäkymmenessä millisekunnissa.[19] Banaanikärpäsellä, kuten todennäköisesti monilla muillakin kärpäsillä, näköhermot ovat myös suoraan yhteydessä siipilihaksiin. Tämä mahdollistaa kärpäsen erittäin nopean reagoinnin liikkeeseen, jopa ilman aivojen väliintuloa.[18]
Näköaisti
[muokkaa | muokkaa wikitekstiä]
Banaanikärpäsen kehittynein aisti on näkö. Noin kahta kolmasosaa banaanikärpäsen aivoista, yhteensä noin 200 000 neuronia, käytetään pelkästään näköaistimusten tulkitsemiseen. Vaikka banaanikärpäsen tilan hahmottamiskyky on huomattavasti huonompi kuin ihmisillä, näköaistimusten käsittelyn nopeus (ajallinen resoluutio) on noin kymmenen kertaa ihmistä nopeampi. Tämä yhdessä lentolihasten ja silmien suoran yhteyden kanssa mahdollistaa petojen tehokkaamman välttämisen ja paremman lennon hallinnan.
Banaanikärpäsen erittäin hyvin kehittyneet verkkosilmät koostuvat 800 pienemmästä silmäyksiköstä, ommatidista. Jokainen ommatidi koostuu kahdeksasta valoreseptorisolusta (nimetty R1–8), niiden tukisoluista, pigmenttisoluista ja verkkokalvosta. Silmien normaali voimakkaan punainen väri johtuu pigmenttisoluista, jotka imevät ylimääräisen sinisen päivänvalon ja siten estävät kärpästä sokaistumasta kirkkaassakaan auringonpaisteessa.[20]
Banaanikärpänen genetiikan tutkimuskohteena
[muokkaa | muokkaa wikitekstiä]Historia
[muokkaa | muokkaa wikitekstiä]- Pääartikkeli: Banaanikärpästutkimuksen historia

Banaanikärpänen on yksi tutkituimmista organismeista ja biologian malliorganismeista. Kalifornian yliopiston perinnöllisyystieteen professori Charles W. Woodworth oli ensimmäinen tutkija, joka kasvatti suurempia määriä banaanikärpäsiä laboratoriossa.[9] Kokeellisen tutkimuksen banaanikärpäsellä aloitti Woodworthin ehdotuksesta Harvardin yliopiston professori William Ernest Castle, jonka oppilas ja työtoveri Frederic Carpenter julkaisi ensimmäisen tieteellisen artikkelin banaanikärpäsestä 1905. Merkittävin 1900-luvun alun banaanikärpästutkijoista oli kuitenkin Columbian yliopiston professori Thomas Hunt Morgan ryhmineen. Hän teki monia kokeita banaanikärpäsillä ja löysi muun muassa 1910 banaanikärpäsen ensimmäisen mutantin, whiten, joka muutti kärpäsen silmät valkoisiksi. Morgan paikallisti mutatoituneen geenin banaanikärpäsen X-kromosomiin, todistaen samalla, että geenit sijaitsevat kromosomeissa. Thomas Hunt Morganille myönnettiin 1933 Nobelin lääketieteen palkinto banaanikärpästen genetiikkaan kohdistuneista tutkimuksista.[9][21]
Ensimmäisen kromosomikartan banaanikärpäselle laati vuonna 1913 Alfred H. Sturtevant, joka työskenteli myös Morganin ryhmässä. T. H. Morgan itse julkaisi ensimmäiset tarkat fysikaaliset paikat useille banaanikärpäsen geeneille vuonna 1938. Banaanikärpäsellä tehdyn varhaisen mutaatiotutkimuksen uranuurtaja 1920-luvulla puolestaan oli Hermann J. Muller, joka indusoi mutaatioita röntgensäteilyn avulla. Keksinnöstään Muller sai lääketieteen Nobelin 1946. Myös evoluutiotutkimusta banaanikärpäsellä tehtiin jo varhain. Yhdysvaltalainen Theodosius Dobzhansky todisti 1930- ja 1940-luvuilla banaanikärpäsen avulla luonnonvalinnan toimivan luonnonpopulaatioissa sekä pystyi selvittämään eri Drosophila-lajien fylogenian.[9]
Banaanikärpäsen suosion syyt
[muokkaa | muokkaa wikitekstiä]Pelkkä sattuma tai tutkimuksen pitkä historia eivät ole syynä banaanikärpäsen suosioon tutkimuskohteena tai sen valintaan tutkimuksen malliorganismiksi. Banaanikärpäsen käyttöön ovat tutkimuksen historian aikana vaikuttaneet useat seikat, kuten:[16][21]
- Lajin pieni koko ja laboratoriossa kasvattamisen helppous
- Lyhyt sukupolven kierto ja suuri sikiävyys
- Pieni kromosomisto
- Sylkirauhasten jättiläismäiset polyteenikromosomit mahdollistavat muun muassa geenien aktiivisuuden tutkimisen
- Koirailla ei ole geneettistä tekijäinvaihduntaa, mikä helpottaa geneettisiä tutkimuksia
- Useita tunnettuja mutaatioita, ja uusien mutaatioiden tuottaminen eli indusointi on helppoa
Nykyinen tutkimus
[muokkaa | muokkaa wikitekstiä]- Pääartikkeli: Banaanikärpäsen nykyinen tutkimus
Jo olemassa olevan tiedon ja tutkimuksen valtava määrä jouduttaa myös uutta tutkimusta. Banaanikärpäsellä tehdään nykyään tutkimusta lähes kaikilla biologian osa-alueilla. Viime aikojen suurimpia saavutuksia banaanikärpästutkimuksessa on ollut sen koko genomin sekvensointi vuonna 2000.[22][23] Samoin monien uusien geenimuuntelutekniikoiden kehitys ja mahdollisuus hankkia useimpia tunnettuja mutanttikantoja tutkittavaksi Internetin välityksellä on helpottanut tutkimusta huomattavasti. Nykyään banaanikärpästä käytetään tutkimuskohteena yhtä lailla kehitysbiologiassa, geenien vuorovaikutusten tutkimuksessa, fysiologiassa kuin ihmisten tautien mallintamisessakin. Banaanikärpäsen käyttöä erityisesti ihmisen sairauksien tutkimuksessa puoltaa varsinkin mahdollisuus tehdä erittäin monenlaisia geneettisiä kokeita. Muun muassa kontrolloitujen risteytysten sekä poistogeenisten yksilöiden tuottaminen on banaanikärpäsellä huomattavan helppoa ja lyhyen sukupolvenkierron vuoksi kokeiden tulokset ovat nopeasti nähtävissä.
Esimerkiksi alkoholin vaikutusten ja alkoholismin tutkimuksessa banaanikärpästä on käytetty erittäin paljon. Banaanikärpästen ja nisäkkäiden alkoholivaste on erittäin samankaltainen: esimerkiksi kärpästen liikkeiden kontrolli heikkenee jo pienen alkoholialtistuksen vaikutuksesta ja annosta lisättäessä ne menettävät liikuntakykynsä täysin. Pitkäaikaisessa altistuksessa kärpäsille kehittyy myös toleranssia alkoholia vastaan. Alkoholinsietokyvyltään erilaisten kärpäskantojen geneettinen tutkimus onkin paljastanut useita alkoholiherkkyyteen vaikuttavia geenejä, joille on löydettävissä vastingeenit myös ihmisestä. Siten kärpäsellä tehtävä tutkimus voi paljastaa yksityiskohtia myös ihmisten alkoholismin perinnöllisyydestä.[24]
Alkoholitutkimuksen lisäksi myös muistin mekanismien selvittämisessä banaanikärpänen on osoittautunut oivalliseksi tutkimuskohteeksi. Sen on osoitettu pystyvän oppimaan monenlaisia asioita, kuten erilaisia käyttäytymismalleja. Banaanikärpäseltä onkin onnistuttu löytämään useita tärkeitä oppimiseen ja muistiin vaikuttavia geenejä. Niiden selvittämiseen on käytetty useita erilaisia geneettisiä tutkimusmenetelmiä, kuten esimerkiksi geenien hiljentämistä ja DNA-mikrosirutekniikkaa. Banaanikärpäsellä tehdyissä pavlovilaisissa käyttäytymiskokeissa puolestaan on pystytty eristämään oppimisvaikeuksellisia kärpäskantoja yhdistämällä tietyt tuoksut sähköshokkiin. Näistä eristetyt mutatoituneet geenit (esimerkiksi dunce, rutabaga ja amnesiac) ovat osoittautuneet tärkeiksi muistin kehityksessä. Tuoreet tutkimukset ovat myös paljastaneet banaanikärpäsen pitkäaikaiseen muistiin vaikuttavia geenejä. Eräiden tutkijoiden mukaan banaanikärpäsen muisti- ja oppimistutkimuksessa ollaankin lähitulevaisuudessa pääsemässä vaiheeseen, jossa voidaan siirtyä yksittäisten geenien tutkimisesta jopa koko muistin kehityksen vuorovaikutusverkoston tutkimukseen.[25][26]
Banaanikärpäsen genomi
[muokkaa | muokkaa wikitekstiä]Kromosomisto
[muokkaa | muokkaa wikitekstiä]
Banaanikärpäsen kromosomisto on pieni: sen diploidinen kromosomiluku 2n = 8. Kromosomipareista kolme on autosomeja ja yksi on sukupuolikromosomipari (X ja Y). Historiallisista syistä banaanikärpäsen X-kromosomia merkitään usein numerolla 1, ja autosomit ovat kromosomit 2–4. Neljäs kromosomi jätetään usein huomiotta sen pienen koon takia, sillä siinä on geenejä sisältävää eukromatiinia vain noin miljoona emäsparia.[22] Geenejä neljännessä kromosomissa on alle sata.[27] Kärpäsen sukupuoli määräytyy X-kromosomien ja autosomien suhteen perusteella, eikä Y-kromosomilla ole siihen vaikutusta toisin kuin esimerkiksi ihmisellä. Jos siis X-kromosomeja on yksi, X-kromosomien ja autosomien suhde on 1:6, jolloin syntyy koiras. Jos X-kromosomeja on kaksi, suhde on 2:6, jolloin syntyy naaras.[28] Banaanikärpäsen Y-kromosomi on myös lähes kokonaan heterokromatiinia.[22]
Genomin koko
[muokkaa | muokkaa wikitekstiä]Drosophilan genomi on noin 180 miljoonan emäsparin (Mb) suuruinen. Genomista kolmasosa on heterokromatiinia ja loput noin 120 Mb eukromatiinia.[22] Vaikuttaa siltä, että noin 60 prosenttia kärpäsen genomista ei sisällä proteiineja koodaavia geenejä.[29] Tällä hetkellä genomin arvioidaan sisältävän 14 601 geeniä.[27] Koko genomi on sekvensoitu vuonna 2000, ja suurin osa siitä on sen jälkeen myös varustettu selityksin eri alueiden biologisesta toiminnasta (engl. annotated).[22] Geneettisesti banaanikärpänen on noin 60–70 -prosenttisesti ihmisen kaltainen, ja vajaalle 3 000 banaanikärpäsen geenille on löydettävissä todennäköisesti samaa alkuperää oleva vastingeeni ihmisestä.[21][30] Noin 77 prosentille ihmisen geeneistä, joiden mutaatioiden tiedetään olevan jonkin sairauden syynä, on löydettävissä homologinen vastine banaanikärpäsen genomista.[31] Siten banaanikärpästä voidaan käyttää vertailevassa genomiikassa myös ihmisen ja muiden nisäkkäiden geenitoiminnan tutkimiseen.
Polyteenikromosomit
[muokkaa | muokkaa wikitekstiä]- Pääartikkeli: Polyteenikromosomi

Banaanikärpäsen kromosomiston erikoisimpia rakenteita ovat sen toukan sylkirauhasten polyteenikromosomit. Ne ovat kaapelimaisia useaan kertaan monistuneita jättiläiskromosomeja. Polyteenikromosomeja tavataan joillakin Diptera-lajeilla kudoksissa, jotka eivät erilaistu aikuisen hyönteisen rakenteiksi. Ne ovat pääosin geenitoimintaa tehostavia rakenteita: useiden identtisten geenikopioiden ansiosta mRNA-synteesi on polyteenikromosomillisissa soluissa erittäin tehokasta. Polyteenikromosomeja tavataankin siksi useimmiten sylkirauhasten solujen kaltaisissa solutyypeissä, joissa vaaditaan paljon jotain tiettyä proteiinia.
Kromosomien koko on erittäin suuri tavallisiin interfaasikromosomeihin verrattuna. Ne voivat olla paksuudeltaan jopa 15–25 μm ja pituudeltaan 100–250 μm. Polyteenikromosomien rakenteelle on tyypillistä tummien kromomeeri- ja vaaleiden interkromomeeriraitojen vaihtelu, joiden avulla kromosomit voidaan erottaa toisistaan. Juovarakenne syntyy DNA-rihmojen lukumäärän vaihtelusta ja erilaisesta pakkautumisasteesta kromosomin eri osissa. Kromosomit ovat organisoituneet kromosentrin ympärille, joka sisältää niiden sentromeerialueet.[32]
Geenien merkintätapa
[muokkaa | muokkaa wikitekstiä]Banaanikärpäsen geenien, mutaatioiden ja eri kantojen merkintä on hyvin tarkkaan määriteltyä. Nimeämiskäytännöt ja merkintätavat juontavat juurensa jo T. H. Morganin ajalta. Tarkkaan säännelty geneettinen nomenklatuuri helpottaa huomattavasti tutkimuksen tekemistä, sillä se mahdollistaa erilaisten banaanikärpäskantojen yksiselitteisen kuvaamisen.
Banaanikärpäsgenetiikassa geenien nimet ovat yleensä johdettu geenin ensimmäisen havaitun mutantin fenotyypistä. Esimerkiksi white-geenin ensimmäisenä kuvattu mutanttialleeli muuttaa kärpäsen silmät valkoisiksi. Jokaisen geenin normaali- eli villimuotoa merkitään plus-merkillä (+) Geenien nimet merkitään aina kursivoituna. Nimi alkaa pienellä alkukirjaimella, jos em. ensimmäinen tunnettu mutantti on ollut resessiivinen ja isolla, jos se on ollut dominoiva. Jokaisesta geenistä voidaan käyttää myös sen 1–3 -kirjaimista lyhennettä (esimerkiksi white = w; hedgehog = hh tai decapentaplegic = dpp). Geenien tuottamista proteiineista käytetään samoja nimiä, mutta ne kirjoitetaan aina isolla eikä niitä kursivoida (esimerkiksi White, W; Hedgehog, HH tai Decapentaplegic, DPP). Jos geenistä tunnetaan useita mutanttialleeleja, ne merkitään yleensä yläindeksillä. (esimerkiksi wa, ww) Samoin villialleelin merkintää voidaan tarkentaa yläindeksillä (esimerkiksi w+, hh+ tai dpp+).Tämän lisäksi on olemassa lukuisia muita käytäntöjä, jotka mahdollistavat niin monimutkaisten risteytysten kuin myös runsaasti geeni- ja kromosomimutaatioita sisältävien kantojen yksiselitteisen kuvaamisen.[16][33]
Toisinaan jotkut geeneille annetuista nimistä voivat olla varsin erikoisia. Useiden väitöskirjatutkijoiden ja jopa edistyneempienkin banaanikärpästieteilijöiden pyrkimyksenä on nimittäin ollut jo Morganin ajoista lähtien keksiä kuvaamilleen uusille geeneille mahdollisimman "hauska", mutta samalla kuitenkin kuvaava nimi. Tämän pitkän tradition tulosta ovat muun muassa shavenbaby-mutantti, jonka toukat ovat karvattomia ja he-is-not-interested-geeni, joka vaikuttaa koiraiden kosiokäyttäytymiseen.[34]
Pyydystäminen ja kasvatus
[muokkaa | muokkaa wikitekstiä]Pyydystäminen
[muokkaa | muokkaa wikitekstiä]
Koska banaanikärpästen elintavat saavat ne viihtymään erinomaisesti myös ihmisasumuksissa, niiden määrä kodeissa voi joskus kasvaa häiritseväksi. Asuntoihin sisälle tunkeutuvia kärpäsiä pyydystetään rakentamalla tappava kärpäsansa esimerkiksi suppilosta, jonka pohjalla on syöttinä pieni määrä jotakin käynyttä nestettä. Syötti voi olla esimerkiksi olutta, väkiviinaetikkaa tai punaviiniä, joka voi olla etikoitunuttakin. Pyydykseen joutuneet kärpäset eivät pääse suppilosta pois ja hukkuvat sinne. Syöttinesteen joukkoon voi lisätä myös astianpesuainetta. Se alentaa nesteen pintajännitystä ja edesauttaa siten kärpästen hukkumista.[13]
Käyttö lemmikkieläinten ravintona
[muokkaa | muokkaa wikitekstiä]Luonnosta tai kotoa eri tavoin pyydystettyjä tai lemmikkikaupasta ostettuja banaanikärpäsiä käytetään myös esimerkiksi lemmikkisammakoiden ja akvaariokalojen ruokana. Esimerkiksi veden pinnasta ravintonsa hankkiville labyrinttikaloille banaanikärpäset ovat suositeltua ravintoa. Ruokintaan käytetään useimmiten kaupan olevia siivettömiä mutanttikärpäskantoja, sillä ne eivät voi karata lentämällä pois terraariosta tai akvaariosta.[35][36] Banaanikärpäsiä voidaan kasvattaa myös kotona. Kasvatus onnistuu parhaiten erlenmeyer-pullon kaltaisessa lasiastiassa, johon on valettu hiivaa ja fruktoosisokeria sisältävää agaroosi- ja mannaryynihyytelöä.[37]

Kasvatus laboratoriossa
[muokkaa | muokkaa wikitekstiä]- Pääartikkeli: Banaanikärpäsen kasvatus laboratoriossa
Lähes vastaavaa banaanikärpästen kasvatusmenetelmää käytetään myös useimmissa laboratorioissa, tosin useimmiten pulloina käytetään tähän tarkoitukseen erityisesti valmistettuja 200 ml:n vetoisia kasvatuspulloja ja 50 ml:n vetoisia risteytyspulloja.[16] Kantoja ylläpidetään yleensä 18 °C:ssa, jossa sukupolvenkierron pituus on noin 4–5 viikkoa. Risteytykset tehdään useimmiten noin 24 °C:ssa, jolloin kierto on nopeampi eikä tuloksia tarvitse odottaa liian pitkään. Kasvatuspulloissa ylläpidettävät kärpäset siirretään uusiin pulloihin noin puolentoista kuukauden välein, jotta kärpäsille on aina tarjolla tuoretta ravintoa eivätkä mahdolliset homeet pääse leviämään ravintoalustassa.[38]
Banaanikärpäsen tauteja
[muokkaa | muokkaa wikitekstiä]- Pääartikkeli: Banaanikärpäsen taudit
Yksi syy banaanikärpäsen suosioon laboratorio-organismina on myös sen taudin- ja loistenkestävyys. Yleisimmät laboratoriokantojen ongelmista johtuvat joko kasvatusalustassa leviävistä homeista tai punkeista. Viime vuosina useissa laboratorioissa on myös raportoitu esiintyvän pikornaviruksiin kuuluvaa DCV-virusta (engl. Drosophila C Virus), joka tappaa koteloituneita toukkia. Useimpia banaanikärpäsen tauteja torjutaan yksinkertaisesti pitämällä kasvatustilat mahdollisimman puhtaina ja eristämällä kaikki laboratorioon saapuvat uudet kannat.[38]
Lähteet
[muokkaa | muokkaa wikitekstiä]- Adams, M.D., et al.: The genome sequence of Drosophila melanogaster. Science, 2000, 287. vsk, s. 2185–2195. Artikkelin verkkoversio. (pdf)
- Ashburner, M., Golic, K.G. & Hawley, R.S.: Drosophila: A Laboratory Handbook. Cold Spring Harbor Laboratory Press, 2005. ISBN 0-87969-706-7
- Portin, P.: Sata vuotta banaanikärpästutkimusta. Tieteessä tapahtuu, 2005, nro 5, s. 40–41. Artikkelin verkkoversio. (pdf) (Arkistoitu – Internet Archive)
Viitteet
[muokkaa | muokkaa wikitekstiä]- ↑ Meigen JW: Systematische Beschreibung der bekannten europäischen zweiflügeligen Insekten. (Volume 6). Schulz-Wundermann, 1830. Teoksen verkkoversio (pdf).
- ↑ Integrated Taxonomic Information System (ITIS): Drosophila melanogaster (TSN 146290) itis.gov. Viitattu 23.12.2010. (englanniksi)
- ↑ a b c Ashburner, M., Golic, K.G. & Hawley, R.S.: Drosophila: A Laboratory Handbook. Cold Spring Harbor: Cold Spring Harbor Laboratory Press, 2005. ISBN 0-87969-706-7
- ↑ FlyBase Stock List FlyBase. Arkistoitu 9.10.2006. Viitattu 27.10.2006. (englanniksi)
- ↑ Chiang, H.C. & Hodson, A.C.: An analytical study of population growth in Drosophila melanogaster. Ecological Monographs, 1950, 20. vsk, s. 173–206. Artikkelin verkkoversio. (html)
- ↑ Bakker, K.: An analysis of factors which determine success in competition for food among larvae of Drosophila melanogaster. Archives Nederlandaises de Zoologie, 1961, nro 14, s. 200–281.
- ↑ a b Weigmann, K., Klapper, R., Strasser, T., Rickert, C., Technau, G.M., Jäckle, H., Janning, W. & Klämbt, C.: FlyMove – a new way to look at development of Drosophila. Trends in Genetics, 2003, nro 19, s. 310–311. FlyMove-sivusto. (Arkistoitu – Internet Archive)
- ↑ Mattila, J.: Imaginaalilevyt – kurssityöohje.. Genetiikan laboratorio, Turun yliopisto, 2003.
- ↑ a b c d e f g Portin, P.: Sata vuotta banaanikärpästutkimusta. Tieteessä tapahtuu, 2005, nro 5, s. 40–41. Artikkelin verkkoversio. (pdf) (Arkistoitu – Internet Archive)
- ↑ Markow, T.A. & O'Grady, P.M.: Drosophila - A Guide to Species Identification and Use, s. 176. Elsevier, 2006. ISBN 978-0-12-473052-6
- ↑ Jari Kahanpää (toim.): Draft Catalogue of Finnish Flies (Diptera:Brachycera) elisanet.fi. Viitattu 27.10.2006. (englanniksi)
- ↑ a b Lumme, J. & Lakovaara, S.: Seasonality and diapause in Drosophilids, s. 3d:171–220. (Teoksessa: Ashburner, M., Carson, H. L. & Thompson, J. N. (toim.). The Genetics and Biology of Drosophila) Academic Press, 1983. ISBN 0-12-064948-9
- ↑ a b c Jyrki Matikainen: Kysy luonnosta – Mistä tulevat mahlakärpäset? 30.9.2006. Turun Sanomat. Arkistoitu 30.9.2007. Viitattu 30.1.2007.
- ↑ Ylen tiedeuutiset: Makea ja kitkerä ovat kiinteästi ohjelmoituna aivoihin 22.1.2006. YLE. Viitattu 1.2.2007.
- ↑ Pitnick, S.: Investment in testes and the cost of making long sperm in Drosophila. American Naturalist, 1996, nro 148, s. 57–80.
- ↑ a b c d Nokkala C. (toim): Genetiikan perusteiden harjoitukset, teoriamoniste. Turun yliopisto, Genetiikan laboratorio, 1996.
- ↑ Marlina Montano: Sexual Behaviors Leading To Mating Successes And Failures In Adult Male And Female Drosophila melanogaster 2001. The Department of Bioagricultural Sciences and Pest Management at Colorado State University. Arkistoitu 28.10.2007. Viitattu 21.3.2007. (englanniksi)
- ↑ a b Sane, S.P.: The aerodynamics of insect flight. The Journal of Experimental Biology, 2003, 206. vsk, s. 4191–4208. Artikkelin verkkoversio. (pdf)
- ↑ Fry, S.N., Sayaman, R. & Dickinson, M.H.: The Aerodynamics of Free-Flight Maneuvers in Drosophila. Science, 2003, 300. vsk, s. 495–498. Artikkelin verkkoversio. (pdf)
- ↑ Ranganathan, R., Malicki, D.M. & Zuker, C.S.: Signal Transduction in Drosophila Photoreceptors. Annual Review of Neuroscience, 1995, 18. vsk, s. 283–317. Artikkelin verkkoversio. (html)[vanhentunut linkki]
- ↑ a b c Vappu Pitkänen: Banaanikärpänen hallinnut laboratorioita yli sata vuotta. Turun Sanomat, 3.11.2009. Artikkelin verkkoversio.
- ↑ a b c d e Adams, M.D., et al.: The genome sequence of Drosophila melanogaster. Science, 2000, 287. vsk, s. 2185–2195. Artikkelin verkkoversio. (pdf)
- ↑ Trans-NIH Fly Initiative: Drosophila White Paper 2001 U.S. National Institute of Health. Arkistoitu 10.3.2007. Viitattu 31.1.2007. (englanniksi)
- ↑ Heberlein, U., Wolf, F.W., Rothenfluh, A. & Guarnieri, D.J.: Molecular Genetic Analysis of Ethanol Intoxication in Drosophila melanogaster. Integrative & Comparative Biology, 2004, 44. vsk, s. 269–274. Artikkelin verkkoversio. (pdf)
- ↑ Sforza, D.M. & Smith, D.J.: Genetic and Genomic Strategies in Learning and Memory. Current Genomics, 2003, 4. vsk, s. 475–485. Artikkelin verkkoversio. (pdf) (Arkistoitu – Internet Archive)
- ↑ Margulies, C., Tully, T. & Dubnau, J.: Review: Deconstructing Memory in Drosophila. Current Biology, 2005, nro 15, s. R700–R713. Artikkelin verkkoversio. (pdf) (Arkistoitu – Internet Archive)
- ↑ a b FlyBase: Drosophila melanogaster genome release 5.1 notes flybase.bio.indiana.edu. 8.12.2006. Viitattu 2.2.2007. (englanniksi)
- ↑ Slee R. & Bownes M.: Sex determination in Drosophila melanogaster. Quarterly Review of Biology, 1990, 65. vsk, nro 2, s. 175–204. Artikkelin tiivistelmä. (html)
- ↑ Halligan, D.L. & Keightley, P.D.: Ubiquitous selective constraints in the Drosophila genome revealed by a genome-wide interspecies comparison. Genome Research, 2006, 16. vsk, nro 7, s. 875–884. Artikkelin tiivistelmä. (html)
- ↑ Venter J. C. et al.: The sequence of the human genome. Science, 2001, 291. vsk, s. 1304–1351. Artikkelin verkkoversio. (pdf)
- ↑ Reiter, L.T., Potocki, L., Chien, S., Gribskov, M. & Bier, E.: A systematic analysis of human disease-associated gene sequences in Drosophila melanogaster. Genome Research, 2001, 6. vsk, nro 11, s. 1114–1125. Artikkelin verkkoversio. (pdf)
- ↑ Griffiths, A. J. F., Miller, J. H., Suzuki, D. T., Lewontin, R. C. & Gelbart, W. M.: An Introduction to Genetic Analysis, 7th Edition. W. H. Freeman, 1999. ISBN 0-7167-3520-2
- ↑ Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K. & Walter, P.: Molecular Biology of the Cell, 4th edition. Garland Science, 2002. ISBN 0-8153-4072-9 Teoksen verkkoversio (html).
- ↑ Vincent, J-P & Salecker, I: Why flies are relevant to medical research (Alkuperäinen julkaisu Mill Hill Essays 2004, ISBN 0-9546302-2-X) 2004. National Institute for Medical Research, London. Arkistoitu 31.12.2007. Viitattu 4.7.2007. (englanniksi)
- ↑ Niina & Joonas Gustafsson: Kiinankellosammakon hoito terraariossa sammakkolampi.net. Arkistoitu 12.7.2007. Viitattu 30.1.2007.
- ↑ Jukka Järvi: Akvaariokalojen elävä ruoka aquahoito.info. Viitattu 1.2.2007.
- ↑ Ville Kivisalmi: Banaanikärpästen kasvatus 2006. Tampereen akvaarioseura. Viitattu 11.4.2007.
- ↑ a b Ashburner, M. & Roote, J.: Maintenance of a Drosophila Laboratory: General Procedures 2007. CSH Protocols. Viitattu 27.6.2007. (englanniksi)
Aiheesta muualla
[muokkaa | muokkaa wikitekstiä]- The WWW Virtual Library: Drosophila – Portaali erilaisiin verkossa oleviin banaanikärpässivustoihin (englanniksi)
- Introduction to Drosophila – Lyhyt ja yksinkertainen banaanikärpäsen ja sen tutkimuksen esittely (englanniksi)
- FlyBase – Kattava tietopankki banaanikärpäsen ja sen sukulaislajien genetiikasta ja molekyylibiologiasta (englanniksi)
- The Interactive Fly (Arkistoitu – Internet Archive) – Tietoa ja kuvia banaanikärpäsen kehityksestä ja genetiikasta (englanniksi)
- Drosophila White Paper (Arkistoitu – Internet Archive) – Perustelut D. melanogasterin genomin sekvensoinnin tärkeydelle (englanniksi)
- Keeping and Breeding Fruit Flies (Arkistoitu – Internet Archive) – Ohjeita banaanikärpästen kasvattamiseen kotioloissa (englanniksi)
- Maintenance of a Drosophila Laboratory: General Procedures – Banaanikärpästen laboratoriokasvatusohjeet ja tietoa niiden taudeista (englanniksi)
- FlyMove (Arkistoitu – Internet Archive) – valtava määrä kuvia ja videoita banaanikärpäsen kehityksestä (englanniksi)
- FlyNome, a database of Drosophila nomenclature (Arkistoitu – Internet Archive) – Banaanikärpäsen geenien nimien etymologiaa ja selityksiä (englanniksi)