Beringia viime jääkaudella
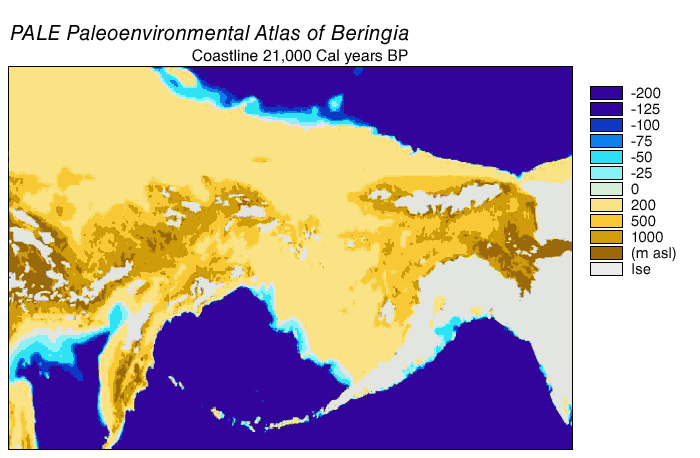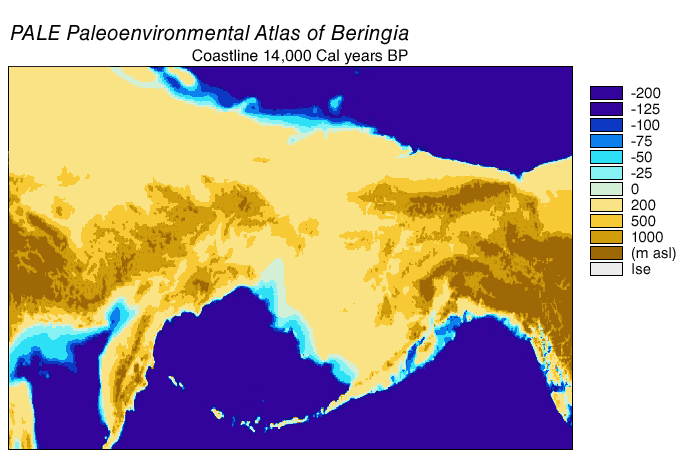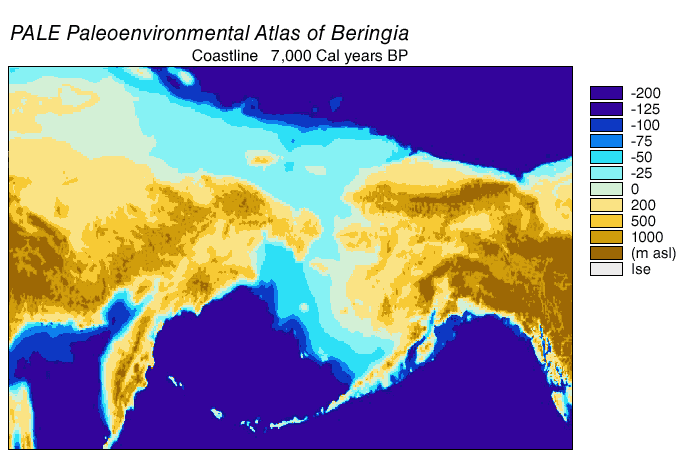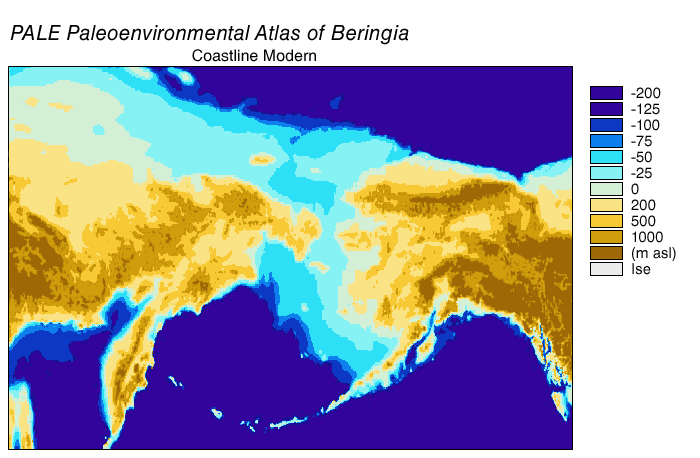
Beringia viime jääkaudella oli Tšuktšien niemimaan ja Alaskan alue sekä näiden välinen alue noin 115 000–11 500 vuotta sitten. Viime jääkaudella Beringiassa eli muun muassa mammutteja, biisoneja ja hevosia, jotka katosivat jääkauden lopussa tai sen jälkeisellä holoseenilla. Jääkauden huippukautena laaja Beringinmeren alue oli kuivaa, alavaa maata. Pleistoseenin jääkausien aikana meri on laskenut ajoittain ja yhdistänyt mantereet Beringinsalmen kohdalla. Beringia oli kuivaa maata viime jääkauden kylminä kausina 10 000–20 000 vuotta kerrallaan kylmyyshuippujen molemmin puolin. Viime jääkauden lopussa 15000-11500 kalenterivuotta sitten Beringiankin ilmasto lämpeni ja kostui nopeasti, jolloin ensin pensaat, sitten puut levisivät aiempien arojen paikalle. Samoihin aikoihin kivikauden metsästäjäihmiset saapuivat ja jääkautinen suureläimistö katosi.
Beringia jääkauden huippukaudella
Beringia viime jääkauden huippukaudella oli Luoteis-Siperian-Alaskan alue viime jääkauden kylmimpänä jaksona, joka ajoitetaan suunnilleen aikaan 24000-17000 radiohiilivuotta sitten. Silloin Itä-Beringiassa kasvoi laajoilla alueilla kuivaa ja puolikuivaa kylmää arotundraa, jota luonnehtivat savikat, maruna, heinät ja ruohot. Kesälämpötila laski laajoilla alueilla korkeintaan 5 astetta. Joidenkin arvioiden mukaan Beringian jokilaaksoissa saattoi olla jopa nykyistä lämpimämpää, kun kesällä viilentävä merenranta oli kauempana ja toisaalta föhn-ilmiö vaikutti. Suuria eläimiä eli suotuisilla paikoilla, mm jokilaaksoissa ja näiden liepeillä. Näitä olivat mammutit, biisonit ja hevoset. Jääkauden kylmimpänä vaiheena yleistyivät eräiden arvioiden mukaan hevoset mammuttien sijaan. Hevoset söivät enemmän kuivassa viihtyviä C4-kasveja kuin mammutit. Läntinen Venäjän puoleinen Beringia oli jääkauden huippukaudella kylmempi varsinkin talvella, mutta sielläkin kasvoi jokilaaksoissa puita.
Beringiassa oli viime jääkauden huippukaudella varsinkin Alaskassa laajoja jäätiköitä ja lössialueita, mutta myös paljon jäätöntä aluetta. Merivirrat kulkivat jääkaudella eri tavoin kuin nyt. Beringian maakannas katkaisi kylmän Jäämereltä tulevan merivirran kulun pohjoiselle Tyynellemerelle. Tämän vuoksi kannas ei pohjoisesta sijainnistaan huolimatta ollut koko leveydeltään jäätikön peittämä, vaikka muualla sekä Euroopassa että Amerikassa jäätiköt ulottuivat paljon etelämmäksikin.
Alaskan jäätiköt olivat lähinnä Kordillieereilla, Aleuteilla ja Brooksin vuorilla[1]. Lumiraja oli Beringiassa noin 250-1500 m korkeudessa, esim. Indigirkan laaksossa 750-1500 m korkeudessa[2]. Ikirouta kattoi koko Beringian[3].
Beringiassa vallitsi jääkaudella jäättömillä alueilla kylminä kausina monin paikoin avoin kuivatundrakasvillisuus. Oli sekä aron että tundran kasveja[4]. Tätä tundraa voidaan kutsua heinä- tai ruohotundraksi[5][6]. Silloin kasvoi Beringiassa heiniä, saroja, marunaa, vaivaispajua ja erilaisia yrtti- eli ruohokasveja[7]. Suuri osa alueesta oli polaariaavikkoa ja sellaisena ihmisille asumiskelvotonta, lukuun ottamatta meren lämmittämää ja kostuttamaa eteläosaa, jossa kasvoi hieman lämpimämpää tundraa[8] ja myös joitain jokilaaksoja. Beringiassa ei liene asuttu viime jääkauden ollessa kylmimmillään. Hopkinskin mukaan Yukon Flatsissa kasvoi metsiä jääkauden kylmimpinä aikoina[9] samoin kuin Indigirka- ja Kolyma-joellakin.
Vuotuisen keskilämpötilan lasku oli Alaskassa maltillinen moniin muihin alueisiin verrattuna[10]. Alaskan keskilämpötila putosi vain noin 4-15 °C[11], mutta Länsi-Beringian eli Luoteis-Siperian lämpötila laski laajoilla alueilla 10-18 °C. Myös ECHAM+BIOME simulaatio sulkee pois ajatuksen, että Beringiassa ei olisi ollut koko alaltaan hyvin kylmää polaariaavikkoa[12].
Erään tietokonemallin mukaan Alaskan jäättömän osan tammikuun keskilämpötila olisi ollut laajoilla alueilla yli -25 °C, muttei yli -10 °C[13][14]. Luoteis-Siperiassa tammikuun keskilämpötila oli elle -25 °C, jopa alle -50 °C. Heinäkuun keskilämpötila oli laajalla alueella 5 °C, mutta alle 10 °C ja Kamtshatkan itäpuolinen meri tätä kylmempi[13]. Luoteis-Siperiassa oli kylmempää, 0-5 °C. Toinen malli antaa Alaskan heinäkuun keskilämpötilaksi laajoilla alueilla yli 6 °C[15], jäätiköiden lähellä +3 °C ja samansuuntaisia lukemia Kolymajoen seuduille.
Erään surviaissääskianalyysin mukaan LGM:n aikana kesälämpötila olisi ollut Länsi-Alaskassa vain noin 2,3-3,5 °C nykyistä alempi mm. Zagoskin Lakessa Saint Michaelin saarella[16] Toiset hyönteistutkimukset väittävät Alaskan heinäkuun lämpötilan pudonneen jääkaududen huippuvaiheessa vain 1-4 °C[16][17].
Erään tietokonemallin mukaan laajoilla alueilla Beringiassa ja jäättömiksi jääneillä Arktisilla saarilla tammikuun keskilämpötila ei laskenut lainkaan tai yleensä 2,5 C[18] ja Baringian keskustan nyt vaden alla olevan alueen talvilämpötila jopa nousi ehkä 2,5 astetta. Joka tapauksessa monien mallien ja kaivausten mukaan Alaskan kesälämpötila nousi LGM:ssä +1 °C - laski -4 °C[16].
Kaikki merkit eivät viittaa siihen, että Beringian lämpötila olisi jääkaudella laskenut. Joidenkin tietojen mukaan Beringiassa olisi viime jääkauden huippukaudella kasvanut Kolyma- joen keskijuoksun altaassa ja Yukon Flatsissa ja Läheisessä Tananajoen laaksossa metsiä[19]. Metsiä olis kasvanut lännempänä myös Kolyma- ja Indigarkajokien laaksossa[20], ja luultavasti myös Lena-joen varrella. Kobuk-joen varrelta Epigurukista on löydetty joenvarsipajustoa jääkauden huippukaudelta[21] ajoitetuna aikoihin 21020-19900 BP[22].
Monilla Luoteis-Siperian alueilla lämpötila ei laskenut jääkaudella niin paljoa kuin Euroopassa. On jopa merkkejä siitä, että ainakin maanpinnan kesälämpötila olisi jopa noussut "kylmänä aikana" muun muassa Kolyma-joen altaassa 1,5-4,5 °C[23]. Tämä on saattanut johtua kosteuden ja pilvisyyden vähenemisestä näillä alueilla.
CCSM3-simulaation mukaan Alaskan puoleisen Beringian lämpötila olisi noussut[16] yli +1,5 °C, joillain jokilaaksoalueilla +4-+10 °C, vaikka kaikkialla muualla maapallolla lämpötila laski, Siperiassa tyypillisesti 4-10 °C myös pohjoisella Kolymajoella[24]. Myös BMRC-simulaatio väittää ainakin joidenkin alaskan osien keskilämpötilan nousseen 2 C[25]. Helmikuun keskilämpötila laski eliöstötutkimusten mukaan noin 12-14 astetta[26] ja vuotuinen keskilämpötila 10 - 8 astetta nykyisestä[27].
Jääkauden kylmän huippuvaiheen aikaan noin 22000-21000 radiohiilivuotta sitten on löydetty Kanadasta ja Alakasta mammutteja, jotka elivät melko lähellä jäätikön reunaa[28]. Näitä paikkoja olivat esim Mackenzie-joen suiston Tunuk Kanadan Luoteisterritorioissa ja Alaskan Bluefish Caves Yukonissa. Todella arktisia olivat Arktisten saarten Ballast Brook, Banksinsaari, Cape James Ross area ja Melvillensaari.[28]
Beringia ja Arctidakin lienevät olleen laajalti asumiskelpoisia viime jääkauden huippukaudellakin[29]. Alaskassa oli jääkauden huippuvaiheessa laajoja lössialueita[30] ja ikiroudpaksuus oli ainakin 600 m, mutta alle 1000 m[31]. Sademäärä laski nykyisestä 100-250 mm[32].
Beringian jääkautinen ekosysteemi
Beringian jääkauden huippukauden aikaisesta luonnosta on kiistelty paljon. Tämä johtuu siitä, että siellä eli monia suuria eläimiä[33] melko kylmässä ja kuivassa ympäristössä, jollainen ei yleensä tuota rehevää kasvustoa. Toisaalta tiedot Beringian kasvillisuudesta perustuvat laajalti siitepölytutkimuksiin, joissa ei kyetä aina tarkoin kasvilajejakaan tunnistamaan, saati sitten sitä kasvoiko esimerkiksi koivu maassa suikertavana vaivaispensaana vaiko puuna. Jääkauden huippukautiset siitepölyjen pienet määrät viittaavat ainakin paikoin hyvin harvaan kasvillisuuteen[34]. Kasvillisuus oli laajalti periglasiaalista ruoho-yrttipitoista tundraa,[35] tundraa[36] tai arotundraa[37], jossa kasvoi mm. Artemisia, Gramineae, Selaginella ja Lycopodeaceae[38]. LDMH-simulaation mukaan nykyään veden alla olevilla alueilla oli pensastundraa ja luultavasti boreaalista metsääkin[35]. Ruoho-yrttitundra on kuivatundraa, jota luonnehtii maruna Artemisia ja tunturisareke Kobresia, ruohot ja sammalet[39]. Tämän tyyppisellä tundralla esimerkiksi lapinvuokko ja vaivaispaju olivat harvinaisia[39].
Yllättäen siitepölyihin perustuviin spekulaatioihin saatiin tarkennuksia, kun jääkauden huippukautinen maannos löytyi. Tulivuoren purkaus hautasi noin 18000 radiohiilivuotta sitten eli 21500 kalenterivuotta sitten luultavasti keväällä pohjoiselle Sewardin niemimaalle ylämaan tundraniityn, jossa sammalen päällä kasvoi tunturisaraketta, muita sarakasveja ja ruohoja[40][41]. Tunturisarake kasvaa nykyään ohutlumisilla, kuivilla, kalkkipitoisilla, tuulisilla vuoristoilla tai tundrilla. Vaivaispajuja oli harvakseltaan. Tuulen tuoman lössin lannoituksen takia olosuhteisin nähden rehevä kasvillisuus ei ollut häiriintynyt, vaan sammalmatto oli yhtenäinen[40], vaikka maanpinta olikin routimisen takia kumpuinen. Niitty kasvoi kesäisin pinnaltaan sulavan jatkuvan ikiroudan päällä lievästi emäksisessä kalkkipitoisessa vähähumuksisessa maannoksessa[40]. Lapinvuokkoa ei kasvanut, mikä viittaa runsaaseen typen ja fosforin määrään[40] ja vakaaseen maaperään. Myöskään niityllä ei kasvanut juurikaan muualla Itä-Beringiassa siitepölyjen mukaan yleistä marunaa, ei vaivaiskoivua, kanervakasveja, suoturvetta. Silttipitoisen lössiperäisen mineraalimaan aktiivinen kerros oli 30-64 cm paksu eri löytöpaikoissa, ja niitty on yrttitundraa, ei aroa[41]. Maannos viittaa ainakin ajoittaiseen märkyyteen vuodenaikojen vaihdellessa[41].
Tämä ylämaan löytö ei kerro mitään esimerkiksi Alaskan sisäosien jokilaaksojen kasvillisuudesta, joiden ajatellaan usein olleen monien muiden jokilaaksojen takia kuivaa aroa, tai myöskään jääkautisesta jokivarsikasvillisuudesta, jonka ajatellaan huippukaudellakin muistuttaneen nykyistä pajuineen ja soineen.
Zimovin vuonna 1995 julkaisemien näkemysten mukaan arotundra erosi tavallisesta tundrasta siten, että kun ilmasto oli kuiva, maannos oli kuiva ja hapekas. Tällöin kivennäisiä irtosi nopeasti rapautuvassa maannoksessa, mikä tarjosi kasveile paljon ravinteita[42]. Tämä mahdollisti kosteata tundraa suuremman biomassan tuotannon, mikä elätti suuria ruohonsyöjäeläimiä, jotka tuottivat kasveille ravinteita kuollessaan, ulostaessaan ja virtsatessaan[42]. Eläimet mm. biisonit sorkillaan myös mylläsivät kulkiessaan maata. Maaperää kuivatti myös runsaan kasviston runsas haihdutus[42]. Guthrien mukaan Beringian arotundra muistutti jääkaudella satavan lössin takia laajalti enemmän ei-hapanta kosteaa tundraa, kuin nykyistä hapanta kosteaa tundraa[43]. Ei-happamassa kosteassa dundrassa eli MNT:ssä maannos oli kalsiumrikas, hienojakoinen ja melko hapan. Runsasravinteinen MNT kasvaa enemmän myrkyttömiä, kalsium- ja muu ravintopitoisia kasveja[43].
Oli kasvillisuus mikä tahansa, Beringiassa viihtyi tyypillinen mammuttifauna[44], joka söi enimmäkseen heinä- ja sarakasveja. Alaskalle tyypillisesti kamelieläimiä[45]
Beringiassa eli jääkaudella mm. arobiisoni, mammutti, kamelilaji Camelops hesternus, hevonen Equus lambei, saiga-antilooppi, vähemmän nykyistä kylmään sopeutunut myskhärkälaji Bootherium bombifrons, wapiti ja hirvi. Saalistajia olivat susi, leijona (P. Atrox, joka oli ehkä jaguaarin sukulainen, ei leijonan), sapelihammastiikeri (Homotherium) ja lyhytnaamakarhu (Arctodus simus). Beringiassa elivät jääkaudella myös jättiläismaalaiskiainen (Megalonyx jeffersoni) ja jättiläismajava (Castoroides ohioensis)[46]. Beringiassa ei ollut villasarvikuonoja[47]. Saiga ja jakkihärkä kuolivat Beringiassa viime jääkauden aikana[47]. Alaskan Beringiassa eli viime jääkaudella myös metsäjäniksiä, tunturisopuleja, punakettuja, kärppiä, vuorikauriita ja naaleja[47].
Eri ilmastosimulaatiot antavat erilaisia tuloksia, siitä mikä olisi ollut yleisin tundran laji. Beringian jääkautiset löydöt viittaavat keskiarktiseen tundraan, jossa heinäkuun keskilämpötila oli 2-6 C niin kuin nyt Kanadan pohjoispuolisilla Banksin ja Victorian saarilla[48].
Länsi-Beringian Berelekin terassien kerrostumissa näkyy 13700 radiohiilivuotta sitten arotundralle tyypillinen heinäkasveja, saroja, marunoita, kohokkikasveja ja myös huomattavasti koivua (Betula exilis) ja pajua[49].
Noin 24 000–17 000 radiohiilivuotta sitten oli hyvin kylmä vaihe viime jääkaudella. Huipussaan kylmyys ja kuivuus olivat noin 22 000–19 000 radiohiilivuotta sitten. Jääkauden kylmimpänäkin kautena uskotaan kasvaneen puita Beringian jokilaaksoissa.lähde? Maruna, savikka, ruohot ja heinät, sarat ja sarakekasvit olivat yleisiä. Kesälämpötila laski monilla alueilla korkeintaan 5 astetta.lähde? Mammutit, biisonit ja hevoset elivät yhä Beringiassa, luultavasti biisoni harvinaistui. Jääkauden kylmimpänä vaiheena yleistyivät eräiden arvioiden mukaan hevoset mammuttien sijaan. Hevoset söivät enemmän kuivassa viihtyviä C4-kasveja kuin mammutit.
Nykyisen näkemyksen mukaan Beringian keskusta oli melko lämmin. Niinpä esimerkiksi Bluefish-luolan lähellä etelään päin olevalla jokirinteellä kasvoi laajalti arotundrakasvillisuutta, varsinaista tundraa oli vasta ylhäällä vuoristossa[50]. Tundrakasvillisuus lisääntyi aromaisella alueella ylöspäin mentäessä. Lössinen aromainen alue kasvoi muun muassa nurmikkaa, marunaa, savikkaa ja kohokkia. Jokilaaksossa kasvoi mm. pajua ja saraa. Jokilaaksossa oli kohtalaisen kosteita ruohoniittyä ja soita[50]. Niinpä mamuttiaro olisi kasvanut jokivarren kasvillisuuden ja tundran välisellä alavalla alueella[48].
Nykyiseen tundraan tehtyjen vertailujen mukaan tyypillisesti Beringian keskiosissa jokien viereistä pajupensasaluetta olisi ollut 5%, märkää saramaata 10%, marunaa, saraa ja heinää kasvavia rinteitä/tasankoja 65% ja 20% lapinvuokkoa jne kasvavaa yläarktista mätäs-sammalkesvillisuutta[51]. Eri eläimet viihtyivät eri alueilla, esimerkiksi hirvi Alces jokilaaksojen pajukoissa, mutta peura loivilla saraheinärinteillä ja vuoristossakin[52]. Mammutti eli muilla alueilla paitsi vuoristotundralla[52].
Esimerkiksi mammutti syö joko pensasalueelta tai sara-alueelta ravinnostaan 40&, ja maruna-heinäalueellta 60%[51]. Biisoni syö märältä sara-alueelta 30%, ja kuivalta maruna-sara-heinäalueelta 70%[51]. Hevonen syö märältä alueelta 20%, ja kuivalta 80%[51].
Tanana-joen reunanaltaasta Fairbanksin läheltä tehtyjen löytöjen mukaan[53] ruohonsyöjät biisoni ja hevonen olivat yleisimpiä eläimiä mutta mammutti melko harvinainen, Cripple Creekissä mammuttia oli vain noin 5% yksilöistä mutta neljäsosa arvioidusta eläinbiomassasta[54]. Suurin osa eläinbiomassasta oli ruohonsyöjiä.
Mammutti, karibu olivat harvinaisempia ja myskihärkä sekä saalistajat vielä harvinaisempia[55].
Tanana-joen laakson löytöjen mukaan mammutti ja hevonen viihtyivät parhaiten alle 12 km päässä joesta, mutta biisoneita oli eniten 12-30 km päässä joesta[56]. Fairbanks Creekin löytöpaikassa joka on 300 m korkeudessa ja 30 km päässä laakson pohjasta biisonia oli löydöistä noin 1/3 ja biomassastakin 6/10[54]. Hirvi Alces Alces tai Alces Americanus kattpoi noin 5 % luista alle 6 km päässä laakson pohjasta[54]. Myskihärkä taas viihtyi parhaiten Engineer Creekissä joka on noin 12 km päässä laakson pohjasta[54] Karibu viihtyi sekä kaukana joesta että lähellä jokea[54]
Esimerkiksi sapelihammastiikeri Amerikan mastodonttia ja saiga-antilooppeja ei Tanana-joen neljässä löytöpaikassa esiinny[54].
On paljon mietitty sitä, mikä olisi ollut suureläimistön tiheys Beringiassa. Se on laajoissa mitoissa riippunut kasviston tuottavuudesta, josta on kiistelty. Afrikan savannilla suureläintiheys on noin 31000 kg/km2, mutta tundralla Nelchina Basinissa Alakassa noin 300 kg/km2[57]. Tällöin 6000-kiloinen mammutti olisi vaatinut noin 20 km2. Vertailun vuoksi jääkautiseen ympäristöön nykyään Fairbanksin lähellä elää eniten hirviä ja jossain määrin karibua ja lihansyöjäeläimiä. Villejä lampaita elää myös jonkin verran[58]. Jotkut ovat verranneet Beringian elämistön oloja nykyiseen myskihärän elinympäristöön tundralla, jossa elää 0,8-1,6 myskihärkää neliökilometrillä. Tällöin Beringiassa olisi elänyt 43-60 mammuttia 1000 km2 alalla ja tämänkin mukaan mammutti olisi vaatinut noin 20 km2. Mutta kasviston tuotto saattoi olla alempi, myskihärkätiheys vain 1/4 myskihärkää km2 ja mamutteja elää harvassa, vain noin 15 mammuttia/1000 km2[59] ja yksi mammutti vaatinut melkein 70 km2. Nykyään Alaskassa elää villejä hevosia Delta Riverin ruohomailla[60]. Monien tutkijoiden mukaan Beringian arotundran suureläimet elivät harvassa[61], ja laidunsivat laikuittain tehokkaasti[48].
Eläimistön kehitys jääkauden lopussa
Alaskan hevoset lienevät olleet keskiaasialaista Equus hemionus -tyyppiä[62], mutta niitä kutsutaan yleensä nimellä Equus caballus alaskai. Eräiden tietojen mukaan esimerkiksi myskihärkää ja saigaa ei olisi ollut Beringiassa jääkauden kylmällä jaksolla noin 20 000–18 000 ka[63]. Jääkauden huippukaudella noin 22 000–19 000 radiohiilivuotta sitten hevonen näyttää löydöistä päätellen olleen huomattavasti mammuttia yleisempi[62]. Hevosen koko alkoi pienetä jo noin 25 000 radiohiilivuotta sitten, ellei aiemmin[62]. Mammutti oli erityisen harvinainen noin 21 000 radiohiilivuotta sitten[62], ja hevonen hyvin yleinen noin 21 500ref name="Guthrie_2003_Horse_Body_Size"/> radiohiilivuotta sitten.
Noin 17 500 radiohiilivuotta sitten alkoi lämmetä Sewardin niemimaalla[64] ja sekä hevonen että mammutti harvinaistuivat, mutta olivat taas yleisiä 16 500 radiohiilivuotta sitten[62]. Noin 16 000 radiohiilivuotta sitten alkoi Sewardin niemimaalla näkyä selviä lämpenemisestä johtuneita kasvillisuusmuutoksia. Jääkauden päättymisen kasvillisuusmuutokset olivat asteittaisia esimerkiksi Alaskassa, josta mammutti hävisi laajoilta alueilta 11 500 radiohiilivuotta sitten eli 13300 kalenterivuotta sitten[65] Saint Paulin saarella mammutteja oli vielä 7 900 radiohiilivuotta sitten. Joidenkin tietojen mukaan mammutti ja hevonen olisivat laajalti hävinneet jo lämpimän Böllingin alussa 14700-14600 kalenterivuotta sitten[66], ja biisoni 12300-10000 kalenterivuotta sitten[66][67], mutta viimeinen Alaskan mammutti kuoli 13000 kalenterivuotta sitten[67].
Pleistoseenin jääkauden aikoina talvet olivat ankaria Beringiassa, mutta ohut lumipeite mahdollisti sen, että ruohoa syövät eläimet löysivät tarpeeksi talvisyötävää. Jääkauden ruohoa ja pensaita kasvavat[68] mammuttiarot sisälsivät tarpeeksi ravinteita, joita tuli muun muassa jään reunalta levinneestä lössistä. Lössi oli syntynyt jään kuluttaessa kiveä. Jääkauden ilmasto oli melko kuiva, ja lumikerros oli matala. Paju oli tavallisin pensas. Noin 14 000–13 000 radiohiilivuotta sitten ilmasto kostui Alaskassa, ja kosteampi pensastundra levisi maahan tästä alkaen[69]. Tämä näkyi selvänä lämpenemisenä ja kasvillisuusmuutoksina myös Pohjois-Alaskan burial Lakessa[64]. 13 500–12 000 radiohiilivuotta sitten Beringian alueella ilmaston vaiheittainen kostuminen kasvatti biisonin määrää, ja väliksi 13 000–12 000 radiohiilivuotta sitten hirven määrää vähäksi aikaa. Mammuttien määrä alkoi pienentyä 13 500 radiohiilivuotta sitten, oli alimmillaan 13 000 radiohiilivuotta sitten ja alkoi pian taas nousta saavuttaen uuden huipun 12 500 radiohiilivuotta sitten, jonka jälkeen määrä alkoi taas pienentyä. 13 500–12 000 radiohiilivuotta sitten kasvoivat rajusti jääkauden kylmälle ilmastolle alun perin ominaisten pajun, ruohon, saran ja marunapensaan määrät. 12 500–12 000 radiohiilivuotta sitten Alaskaan saapui ihminen. Ihmisen saapuminen, jääkauden kasvillisuuden pieneneminen ja koivun yleistyminen näyttävät osuvan yksiin.
Hevonen hävisi 12 500 radiohiilivuotta sitten[62] eli noin 14200 kalenterivuotta noin 500 vuotta ennen varmistettua ihmisen saapumista[62] mitä luultavimmin ennen nuoremman dryaskauden kylmenemistä[65][70]. Mammutti oli yleinen vielä 13 000 radiohiilivuotta sitten, mutta harvinaistui tämän jälkeen nopeasti[62]. Guthrien mukaan juuri 13 000–12 500 radiohiilivuotta ilmaston lämpenemin nja kostuminen aiheutti Alaskan metsittymisen ja soistumisen[62]. Pwnsastundra oli 12 000 radiohiilivuotta sitten yleinen Alaskassa[69]. Maaperä happamoitui ja uudet kasvilajit olivat vähäravinteisia ja biomassan tuotantokin väheni[62]. Arotundran korvautuminen pensastundralla ja muulla tundralla haittasi eniten hevosta, joka hävisikin ensin[62]. Ihminen saapui Alaskaan noin 12 000 radiohiilivuotta sitten tai hieman myöhemmin[62].
12 500–11 000 radiohiilivuotta sitten vaivaiskoivu yleistyi rajusti. Noin 12 500–11 500/11 000 radiohiilivuotta sitten jääkausikasvillisuus harvinaistui nopeasti, muttei hävinnyt kokonaan. Noin 12 000 radiohiilivuotta sitten lämpimimmän kuukauden keskilämpötila alkoi nousta, ollen nykyistä suurempi jo 11 000 radiohiilivuotta sitten[69]. Nuorempi dryaskausi viilensi ilmastoa alakassa 10 800–10 000 radiohiilivuotta sitten[69] ja aiheutti ehkä yrttitundran yleistymisen joillain Alaskan alueilla 10 500–10 200 radiohiilivuotta sitten[69]. Noin 11 000 radiohiilivuotta sitten kesä oli hyvin lämmin, ja talvi hyvin kylmä Alaskassa[69]. Noin 11 000–9 000 radiohiilivuotta sitten pohjoiseen ja keskiseen Alaskaan saapui laajalti haapaa[69]. 10 400 radiohiilivuotta sitten näkyi lämpöhuippu[64]. Noin 9 000 radiohiilivuotta sitten Alaskan kasvillisuus oli suunnilleen vakiintunut, vaikka pieniä muutoksia tapahtui yhä[64]. Boreaalinen havumetsä levisi maahan lopulta noin 5 000 radiohiilivuotta sitten holoseenin keskivaiheilla.
Beringian asuttaminen
Ei tiedetä täsmälleen milloin Beringia asutettiin. Tämä lienee tapahtunut Siperiasta käsin. Myöhäispaleoliittiset metsästäjät etenivät Beringian länsipuolisessa Siperiassa pohjoisemmas joskus 15 500–15 000 vuotta sitten. Näin on voinut käydä Beringiassakin. Noin 14 700 vuotta sitten maapallon ilmasto lämpeni rajusti, ja Beringiaan ilmestyi lämmöstä kielivää pensastundraa noin 14 000 vuotta sitten, jolloin luultavasti ihmiset asuttivat Beringian ja sitä kautta pian Amerikan. Noin 13 000–11 000 vuotta Beringian kautta levisi Amerikkaan aiemmin Keski-Aasiassa kehittynyt mikroteriä valmistanut metsästysperinne, joka eri arvioiden mukaan ei olisi ollut sama kuin Amerikan asuttanut vaellusaalto.lähde?
Milloin Beringia oli mannerta
Ei ole täysin täsmällisiä tietoja siitä, milloin Beringia oli kuivaa maata, sillä tiedot viime jääkauden aikaisista meren pinnan vaihteluista ovat sen verran hatarat. Veiksel-jääkauden jäätikkö oli laajimmillaan muun muassa 55 000, 42 000, 30 000 ja 14 000 vuotta sitten, jolloin maasilta oli auki 5 000–10 000 vuotta maksimien molemmin puolin.[71]
Beringia oli maata 115 000–108 000, 95 000–92 000, 78 000–65 000, 52 000–44 000, 35 000/32 000–12 000 vuotta sitten.[72]lähde tarkemmin?. Pessimistisemmän arvion mukaan Beringia olisi ollut maata viimeksi vain 26 000–12 000 vuotta sitten.
Beringian maasillan katoaminen
Beringian maasilta supistui hyvin hitaasti 20 000–15 000 vuotta sitten, jolloin jäätiköt vetäytyivät melko hidasta vauhtia. 15 000 vuotta sitten leveän Beringian maa-ala oli 2 000 000 km², kun merenpinta oli 90 metriä nykyisen alapuolella. Noin 14 000 vuotta eli 12 000 radiohiilivuotta sitten Beringia oli hyvin leveä, ja se alkoi kaveta vauhdilla noin 13 500 vuotta sitten. Beringia supistui nopeimmin veden kohotessa 70–50 metriä nykyisen tason alle.
Noin 13 000 vuotta sitten (11 000 radiohiilivuotta sitten) Beringian silta kapeni nopeasti, merenpinta oli 60 m nykyisen alapuolella ja maa-ala 1,7 miljoonaa km². Lopullisesti Beringia saattoi hävitä jo noin 12 900[67]. –12 400 vuotta sitten. Varmasti viimeistään noin 11 000 vuotta sitten maayhteys meni täysin poikki, kun merenpinta oli 48 metriä nykyisen tason alapuolella ja nykyään veden peitossa olevaa alaa oli kuivana noin 130 000 km².
Noin 7 000 vuotta sitten Beringin alueen rantaviivat olivat hyvin lähellä nykyistä. Noin 6 000 vuotta sitten rannikkoviivat asettuivat nykyiselle paikalleen.
Lähteet
- Beringia in the Cenozoic era, V. S. Kothekar, Editor-in-Chief V.L.Kontrimavichus, 1985, A.A. Balkema Rotterdam 1986, Printed in India Recha Printers Pvt. Ltd, New Delhi-110020Russian Translations Series 28, ISBN 90-6191-445-0, Venäjänkielinen alkuteos Beringiya v Kainozoe, Vladivostok 1976, Translator Dr. R Chakravarty
- V.S. Kothekar: Beringia in the Cenozoic era. {{{Julkaisija}}}, 1985.
- Tage Nilsson: The pleistocene. Reidel, 1983. ISBN 90-277-1466-5.
- Marck: '. . ISBN. }}
- Frederick Hadleigh West: American Beginnings, the Prehistory and Palaecology of Beringia. University of Chicago Press, 1996. ISBN 0-226-89399-5.
- Dale Guthrie: '. . ISBN.
- Sulkakäärme ja jaguaarijumala. Tampereen taidemuseo, 1997. ISBN 952-9549-43-1.
- Beringia in the Cenozoic era, V. S. Kothekar, Editor-in-Chief V.L. Kontrimavichus, 1985, A.A. Balkema Rotterdam 1986, Printed in India Recha Printers Pvt. Ltd, New Delhi-110020 Russian Translations Series 28, ISBN 90-6191-445-0, Venäjänkielinen alkuteos Beringiya v Kainozoe, Vladivostok 1976, Translator Dr. R Chakravarty
- V.S. Kothekar: Beringia in the Cenozoic era. Joku yritys, 1985. ISBN numero tähän.
- Tage Nilsson: The pleistocene. Reidel, 1983. ISBN 90-277-1466-5.
- Merck, John: Musings on Alaska during the last ice age Joku yritys.
- Frederick Hadleigh West: American Beginnings: The Prehistory and Palaeoecology of Beringia. University of Chicago Press, 1996. ISBN 0-226-89399-5.
- Guthrie, R. Dale: Joku kirja. Joku yritys, puuttuu. ISBN numero tähän.
- puuttuu: Sulkakäärme ja jaguaarijumala. Tampereen Taidemuseo, puuttuu. ISBN numero tähän.
- John F Hoffecker, Scott A. Elias: Human ecology of Beringia. Columbia University Press, 2007. ISBN-13 978-0-231-13060-8.
- Atlas of Paleoclimates and Paleoenvironmants of the Northern Hemisphere - Late Pleistocene-Holocene, Editors-in-chief Burkhard Frenzel, Marton Pecsi, Andrej Alekseevich Velichko, Geographical Researh Institute, Hungarian Academy of Sciences, Gustav Fischer Verlag Stuttgart, Budapest-Stuttgart 1992, ISBN 953-7395-24-5 Budapest, ISBN 3-437-30685-X Stuttgart
Viitteet
| Tämän artikkelin tai sen osan viitteitä on pyydetty muotoiltavaksi. Voit auttaa Wikipediaa muotoilemalla viitteet ohjeen mukaisiksi, esimerkiksi siirtämällä linkit viitemallineille. Tarkennus: muotoilua ja samojen yhdistelyä |
- ↑ Nilsson 1983, 366
- ↑ Nilsson 1983, s 312
- ↑ Nilsson 1983, s 314
- ↑ New Spruce (Picea spp.) Macrofossils from Yukon Territory: Implications for Late Pleistocene Refugia in Eastern Beringia G.D. ZAZULA,1,2 A.M. TELKA,3 C.R. HARINGTON,4 C.E. SCHWEGER5 and R.W. MATHEWES1, ARCTIC VOL. 59, NO. 4 (DECEMBER 2006) P. 391–400
- ↑ [1]
- ↑ Late Quaternary Vegetation History of Northern North America Based on Pollen, Macrofossil, and Faunal Remains Arthur S. Dyke Géographie physique et Quaternaire, Volume 59, numéro 2-3, 2005, p. 211-262
- ↑ Pollen Evidence for Late Pleistocene Bering Land Bridge Environments from Norton Sound, Northeastern Bering Sea, Alaska Thomas A. Ager* and R. Lawrence Phillips† Arctic, Antarctic, and Alpine Research 40(3):451-461. 2008 doi: 10.1657/1523-0430(07-076)[AGER]2.0.CO;2
- ↑ Wiik: Mistä Amerikka sai intiaaninsa
- ↑ The Pleistocene, Tage Nilsson, Reidel 1983, Reidel Member of Kluwer Group, Dordrecht, ISBN 90-277-1466-5, s. 408, alkup. lähde Hopkins 1967b The Bering land Bridge, Stanford University Press 1967
- ↑ Frenzel 1992, s. 41.
- ↑ High-resolution climate simulation of the last glacial maximum, Seong-Joong Kim Æ Thomas J. Crowley Æ David J. Erickson Æ Bala Govindasamy Æ Phillip B. Duffy Æ Bang Yong Lee Received: 13 December 2006 / Accepted: 17 October 2007 / Published online: 6 November 2007,Clim Dyn (2008) 31:1–16 DOI 10.1007/s00382-007-0332-z
- ↑ Modelling global terrestrial vegetation-climate interactio] Martin Claussen Viktor Brovkin ... , The Royal Society, 1998
- ↑ a b [polarmet.osu.edu/PolarMet/PMGFulldocs/bromwich_toracinta_jc_2004.pdf Polar MM5 Simulations of the Winter Climate of the Laurentide Ice Sheet at the LGM], DAVID H. BROMWICH, E. RICHARD TORACINTA, HELIN WEI...,Manuscript received 21 November 2003, in final form 4 March 2004
- ↑ HIGH RESOLUTION LGM SIMULATIONS OVER THE LAURENTIDE ICE SHEET USING POLAR MM5
- ↑ LGM SUMMER CLIMATE OF THE LAURENTIDE ICE SHEET SIMULATED BY POLAR MM5 E. Richard Toracinta1, David H. Bromwich1,2, Robert Oglesby3, James Fastook4, and Terence Hughes4
- ↑ a b c d Late Quaternary paleoclimate of western Alaska inferred from fossil chironomids and its relation to vegetation histories : Kurek, J., Cwynar, L. C., Ager, T. A., Abbott, M. B., Edwards, M. E. (2009). Late Quaternary paleoclimate of western Alaska inferred from fossil chironomids and its relation to vegetation histories. Quaternary Science Reviews, 28 (9-10), 799 - 811, doi:10.1016/j.quascirev.2008.12.001
- ↑ Mutual climatic range reconstructions of seasonal temperatures based on Late-Pleistocene fossil beetle assemblages in Eastern Beringia] Scott A Elias,Quaternary Science Reviews Volume 20, Issues 1-3, January 2001, Pages 77-91 Beringian Paleoenvironments - Festschrift in Honour of D.M. Hopkins vailable online 18 December 2000, doi:10.1016/S0277-3791(00)00130-X
- ↑ Modelling past cold climates, Jenny Brandefelt, KTH Mechanics, LANDCLIM workshop in Norrköping, February 23, 2011
- ↑ Tage Nilsson, The Pleistocene, sivu 408 kartta Fig. 16.21, orig lähde Hopkins 1967b ja myös Björn Kurtén, Jääkausi s 139
- ↑ Kurten 1972, s. 141.
- ↑ American Beginnings, Frederick West jne., The University of Chicago Press, 1996, ISBN 0-226-89399-5, s 25
- ↑ West 1996, s 24
- ↑ Tundra-Steppe Assemblages of Insects and the Reconstruction of the Late Pleistocene Climate in the Lower Reaches of the Kolyma River A. V. Alfimov, D. I. Berman, and A. V. Sher, Received September 24, 2002
- ↑ Last Glacial Maximum and Holocene Climate in CCSM3 BETTE L. OTTO-BLIESNER AND ESTHER C. BRADY, JOURNAL OF CLIMATE, VOLUME 19, 7 November 2005.
- ↑ Influence of vegetation changes during the Last Glacial Maximum using the BMRC atmospheric general circulation model, Wyputta, U.; McAvaney, B.J., Climate Dynamics; Sep2001, Vol. 17 Issue 12, p923
- ↑ Frenzel 1992, s 39
- ↑ Frenzel 1992, s 43
- ↑ a b The Eastern Limit of Beringia: Mammoth Remains from Banks and Melville Islands, Northwest Territories C.R. HARINGTON Received 18 January 2005; accepted in revised form 8 April 2005, ARCTIC VOL. 58, NO. 4 (DECEMBER 2005) P. 361– 369
- ↑ Frenzel 1992, s. 63.
- ↑ Frenzel 1992, s 51
- ↑ Frenzel 1992, s. 49.
- ↑ Frenzel 1992, s. 45.
- ↑ Marck 2011
- ↑ Pisaric, M.F.J., MacDonald, G.M., Velichko, A.A. and Cwynar, L.C. 2001. The late-glacial and post-glacial vegetation history of the northwestern limits of Beringia based on pollen, stomate and megafossil evidence. Quaternary Science Reviews. 20:235-245, Kohta s 241
- ↑ a b Climate change and Arctic ecosystems: 2. Modeling, palaeodata-model comparisons, and future projections, J. O. Kaplan, N. H. Bigelow et al, JOURNAL OF GEOPHYSICAL RESEARCH, VOL. 108, NO. D19, 8171, doi:10.1029/2002JD002559, 2003
- ↑ Frenzel 1992, s. 57.
- ↑ Frenzel 1992, s 59
- ↑ Frenzel 1992, s. 55.
- ↑ a b Climate change and Arctic ecosystems: 1. Vegetationchanges north of 55 N between the last glacial maximum, mid-Holocene,and present, Nancy H. Bigelow, Linda B. Brubaker,JOURNAL OF GEOPHYSICAL RESEARCH, VOL. 108, NO. D19, 8170, doi:10.1029/2002JD002558, 2003
- ↑ a b c d Full-glacial upland tundra vegetation preserved under tephra in the Beringia National Park, Seward Peninsula, Alaska Victoria G. Goetcheus, Hilary H. Birks, Quaternary Science Reviews Volume 20, Issues 1–3, January 2001, Pages 135–147
- ↑ a b c [The full-glacial environment of the Northern Seward Peninsula, Alaska, reconstructed from the 21,500-year-old Kitluk paleosol] Hofle, Claudia, Edwards, M.E., Hopkins, D.M., Mann, D.H., and Ping, Chien-Lu, 2000, Quaternary Research v. 53, n. 2., p. 143-153, doi:10.1006/qres.1999.2097
- ↑ a b c Hoffecker%Elias 2007, s 58, s 59, Alkuperäine lähde Zimov et al 1995
- ↑ a b Hoffecker%Elias 2007, s 57
- ↑ Frenzel 1992, s. 61.
- ↑ Frenzel 1992, s 60
- ↑ usings on Alaska during the last ice age John Merck April 26, 2011
- ↑ a b c Kurten 1972, s. 140.
- ↑ a b c PALEOECOLOGICAL IMPLICATIONS OF PLANT/ANIMAL GRAZING RELATIONS ON THE MAMMOTH STEPPE OF EASTERN BERINGIA, Andre Antinori, Nebraska Anthropologist, 1-1-1995 ja Ritchie 1984, (Cwynar and Ritchie 1980, Cwynar 1982)
- ↑ Kothekar 1985, s. 92.
- ↑ a b Grant D. Zazula a, ,1, Charles E. Schweger a, Alwynne B. Beaudoin b, George H. McCourt c: [doi:10.1016/j.quaint.2005.03.010 Macrofossil and pollen evidence for full-glacial steppe within an ecological mosaic along the Bluefish River, eastern Beringia] (PDF) Quaternary International 142–143. 2006. Elsevier. (englanniksi)
- ↑ a b c d Chapter 4: Mammoth Fauna on Arctic Tundra Hans Krause research report
- ↑ a b Palaeoecology of Beringia, David Hopkins et al, Academic 1982, s. 312
- ↑ West 1996, s 121
- ↑ a b c d e f West 1996, s 122
- ↑ West 1996, s 122, s 123
- ↑ West 1996, s 126
- ↑ West 1996, s 124
- ↑ West 1996, s 127
- ↑ Muskoxen and their hunters: a history Tekijät Peter C. Lent, s. 27.
- ↑ West 1996, s. 128.
- ↑ Guthrie
- ↑ a b c d e f g h i j k l R. Dale Guthrie: Rapid body size decline in Alaskan Pleistocene horses before extinction, Nature 426, 169–171 (13 November 2003)doi:10.1038/nature02098
- ↑ The most common fossil mammals of Beringia during the interval of near maximum continental ice cover. Extinct or extirpated taxa are marked with an asterisk Arthur S. Dyke Géographie physique et Quaternaire Volume 59, numéro 2–3, 2005, s. 211–262, Late Quaternary Vegetation History of Northern North America Based on Pollen, Macrofossil, and Faunal Remains
- ↑ a b c d Kurek Cwynar: Late Quaternary paleoclimate of western Alaska inferred from fossil chironomids and its relation to vegetation histories, Quaternary Science Reviews 2009 xxx, ja CalPal-takaisinkalibraatio
- ↑ a b A calendar chronology for Pleistocene mammoth and horse extinction in North America based on Bayesian radiocarbon calibration,Caitlin E. Bucka, and Edouard Bard, doi:10.1016/j.quascirev.2007.06.013 Quaternary Science Reviews, Volume 26, Issues 17–18, September 2007, sivut 2031–2035
- ↑ a b Hoffecker&Elias 2007, s 73
- ↑ a b c Hoffecker&Elias 2007, s 75
- ↑ Mass Extinct Alaska The Why Files 2010, University of Wisconsin, Board of Regents.
- ↑ a b c d e f g Beringian paleoecology: results from the 1997 workshop, Scott A. Elias, Quaternary Science Reviews 20 (2001) 7/13
- ↑ Rapid body size decline in Alaskan Pleistocene horses before extinction, R. Dale Guthrie, Nature 426, 169–171 (13 November 2003) | doi:10.1038/nature02098; Received 4 February 2003; Accepted 23 September 2003
- ↑ Jqjacobs.net
- ↑ Sulkakäärme ja jaguaarijumala